熱線:021-66110810,66110819
手機:13564362870
熱線:021-66110810,66110819
手機:13564362870
2.17.S-硫水合蛋白的測定
采用兩種方法檢測S-硫水合蛋白。為鑒定潛在的S-硫水合蛋白,按照先前描述的方法進行馬來酰亞胺測定。紅細胞在4°C下以250 mM HEPES-NaOH(pH 7.7)、1 mM EDTA和0.1 mM新亞甲藍組成的RIPA-A緩沖液中裂解30分鐘。裂解液以20,000×g離心10分鐘。收集上清液,然后在4°C下與2 mM Alexa Fluor 680標記的C2馬來酰亞胺孵育2小時,期間偶爾輕輕混合,隨后在4°C下與或不與1 mM DTT孵育1小時。裂解液用于SDS-PAGE凝膠電泳。從凝膠中收集具有S-硫水合的蛋白條帶,通過LC-MS/MS分析以鑒定可能的S-硫水合蛋白。為測定Hb和BPGM的S-硫水合,按照先前描述的方法進行生物素交換測定。紅細胞在HEN緩沖液中均質化,該緩沖液由250 mM HEPES-NaOH(pH 7.7)、1 mM EDTA和0.1 mM新亞甲藍組成,并補充了100μM去鐵胺。在4℃條件下以13,000×g離心30分鐘后,將上清液與HEN緩沖液(含2.5%SDS和20 mM甲硫基甲磺酸甲酯)于50℃孵育20分鐘,期間頻繁渦旋。隨后加入丙酮去除甲硫基甲磺酸甲酯,混合液在-20℃下靜置20分鐘使其沉淀。離心后,將蛋白質重懸于含1%SDS的HEN緩沖液(即HEN-S緩沖液)。向懸浮液中加入不含抗壞血酸的二甲基亞砜配制的生物素-HPDP溶液,于2℃條件下孵育3小時。
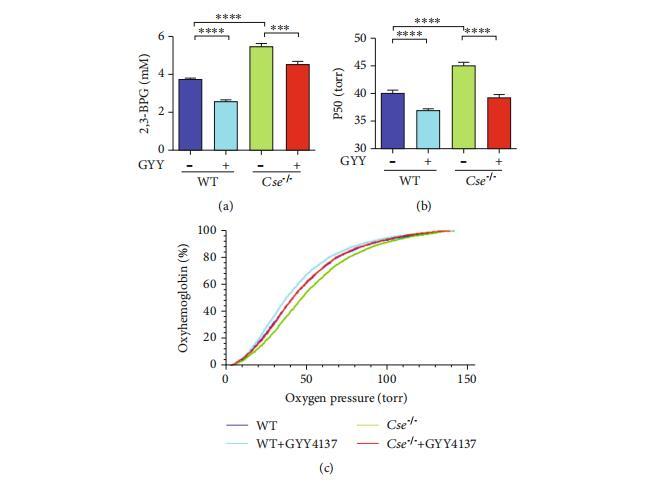
圖1:硫化氫水平降低導致Cse-/-小鼠紅細胞中2,3-BPG生成增加和Hb-O2親和力下降。野生型和Cse-/-小鼠分別給予生理鹽水或GYY4137(133μmol/kg)。(a–c)注射24小時后分離紅細胞以測定2,3-BPG水平(a)和氧解離曲線((b)P50值;(c)氧解離曲線的代表性跡線)。數據以均值±標準誤表示(n=10)。***p<0.001,*****p<0.0001。
2.18統計分析
所有統計分析均使用SPSS 20軟件完成。數據均以均值±標準誤表示。通過Shapiro-Wilk檢驗評估數據的正態分布情況。統計顯著性判定依據樣本分布及方差齊性。兩組間統計比較采用雙尾學生t檢驗。單因素方差分析結合Bonferroni校正。使用條款詳見Wiley在線圖書館的條款與條件;開放獲取文章受適用的知識共享許可協議約束。多組分析采用事后檢驗和Kruskal-Wallis檢驗結合Dunn事后檢驗。統計學顯著性判定標準為p<0.05。
3結果
3.1 Cse?/?小鼠紅細胞中2,3-BPG水平降低且Hb氧親和力增加
我們使用Cse-/-小鼠探索了硫化氫在紅細胞中的功能作用,因為Cse-/-小鼠顯示出顯著降低的硫化氫水平。通過代謝組學技術,我們在紅細胞中鑒定出98種代謝物,并發現與野生型小鼠相比,Cse-/-小鼠的多個代謝途徑(如丙酮酸代謝、糖酵解或糖異生)發生了顯著變化。值得注意的是,與野生型小鼠相比,Cse-/-小鼠紅細胞中的代謝物中2,3-BPG顯著升高。隨后通過分光光度法測定證實了Cse-/-小鼠紅細胞中2,3-BPG水平的升高(圖1(a))。由于2,3-BPG是一種特異性變構調節劑,可調節Hb-O2親和力,我們檢測了Cse-/-小鼠的氧解離曲線。如預期,與野生型小鼠相比,Cse-/-小鼠的50%氧飽和度(P50)值顯著升高了4至6 mmHg(圖1(b)和1(c))。為確保Cse-/-小鼠中2,3-BPG和P50水平的升高是由于硫化氫生成減少所致,我們用緩釋硫化氫供體GYY4137處理了Cse-/-小鼠。如圖1(a)–1(c)所示,GYY4137(133μmol/kg)處理導致2,3-BPG和P50水平降低。
值得注意的是,GYY4137本身降低了野生型小鼠紅細胞中2,3-BPG和P50水平。由于pH是Hb-O2親和力和2,3-BPG形成的主要決定因素之一,我們還檢測了紅細胞中的pH值。在野生型(WT)與Cse-/-小鼠之間未發現pH的顯著差異,這表明Cse-/-小鼠紅細胞中2,3-BPG和P50水平的升高與pH無關。已知CSE在組織中廣泛表達。一些研究表明CSE活性在紅細胞中被識別。小鼠紅細胞能夠產生硫化氫并表達CSE蛋白。CSE-/-小鼠的硫化氫生成速率顯著降低。事實上,外周組織和血清產生的硫化氫可擴散至紅細胞。因此,我們進行了野生型與Cse-/-小鼠之間的相互骨髓移植(BMT)。通過BMT生成了三組小鼠:(1)“WTto-Cse-/-”組通過將野生型小鼠骨髓移植至Cse-/-小鼠,以確定除骨髓來源細胞外,外周組織中CSE的缺乏是否會影響野生型紅細胞對氧合血紅蛋白(HbO2)的親和力;(2)“Cse-/--to-WT”組通過將Cse-/-小鼠骨髓移植至野生型小鼠,以檢查僅在骨髓來源細胞中CSE的缺乏是否足以影響Hb-O2親和力;(3)“WT-to-WT”組為野生型小鼠骨髓移植至野生型小鼠。如預期,“WTto-Cse-/-”小鼠表現出顯著差異。
與“WT-to-WT”和“Cse-/--to-WT”小鼠相比,循環系統中的硫化氫水平較低(圖2(a))。相應地,“WT-to-Cse-/--”小鼠的紅細胞2,3-BPG和P50水平顯著高于“WT-to-WT”和“Cse-/--to-WT”小鼠(圖2(b)和2(c))。GYY4137處理可增加“WT-to-Cse-/--”小鼠循環中的硫化氫水平(圖2(a))。“WT-to-Cse-/--”小鼠中升高的紅細胞2,3-BPG和P50水平被GYY4137逆轉(圖2(b)和2(c))。這些數據表明,除骨髓來源細胞外,外周組織和血清產生的硫化氫對2,3-BPG的生成具有抑制作用,從而維持紅細胞中的Hb-O2親和力。Cse-/--小鼠表現出高血壓,因為硫化氫通過控制KATP通道活性是一種重要的血管擴張劑。
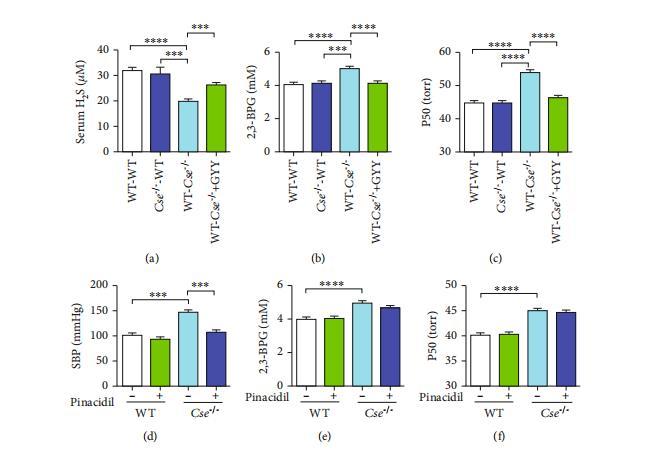
圖2:2,3-BPG和P50的水平取決于外周組織產生的硫化氫,但與Cse-/-小鼠的血壓無關。(a–c)在野生型和Cse-/-小鼠之間進行了骨髓移植。骨髓移植8周后,“野生型-野生型”和“Cse-/---野生型”小鼠接受了生理鹽水,而“野生型-Cse-/--”小鼠則接受了生理鹽水或GYY4137(133μmol/kg)。治療24小時后檢測了循環硫化氫水平(a)、紅細胞2,3-BPG水平(b)和P50(c)。(d–f)野生型和Cse-/--小鼠接受了匹那西地(2.8μmol/kg)或生理鹽水治療。治療24小時后檢測了血壓(d)、紅細胞2,3-BPG水平(e)和P50(f)。數據以均值±標準誤表示(n=8)。
 相關新聞
相關新聞