熱線:021-66110810,66110819
手機:13564362870
熱線:021-66110810,66110819
手機:13564362870
結果
根際沉積物理化指標
N2近根際子隔室中的NO?-N濃度最高(1.46±1.66 mg l?1),根區和非根際區最低(分別為0.30±0.14和0.39±0.16 mg l?1)(F=3.611, P=0.003)(表1)。NH?-N濃度(10.61±7.80 至 13.28±9.49 mg l?1)(F=0.169, P=0.984)和pH(7.56±0.23 至 7.62±0.14)(F=0.151, P=0.962)在各根箱隔室之間無顯著變化。氧滲透深度(Dop)在各隔室間從14.4±6.6降至10.6±4.5 mm,但這并不顯著(F=1.136, P=0.349)。

表1 沉水植物菹草根際和非根際沉積物及孔隙水的理化性質。數值為平均值(標準差);n=12。
注:R和Non分別代表根區隔室和非根際區隔室的沉積物樣品。N1-N5代表取自近根際區1至5毫米子隔室的沉積物樣品。
- 表示根系阻礙了根區隔室內的微電極測量。
- 表示未測量非根際區。
同一列數值后不同字母表示存在顯著差異(P < 0.05)。
類產氧甲烷桿菌16S rRNA基因多樣性
本研究共獲得324條NC10門細菌序列(表2)。所有序列根據97%相似性劃分為16個OTUs。每個根箱夾層檢測到約45-50條序列,代表4到6個不同的OTUs(表2)。在N1近根際子隔室中發現了最多的OTUs(6個),而在根區(R)和N5近根際子隔室中發現了最少的OTUs(各4個)。文庫覆蓋率值在0.94至1.00之間,表明菹草根際沉積物中的類產氧甲烷桿菌16S rRNA基因序列在這些克隆文庫中已充分代表。N2近根際子隔室具有最高的多樣性,香農指數和Chao1豐富度估計量分別為1.16和5。在N3近根際子隔室觀察到最低的多樣性,香農指數和Chao1豐富度估計量分別為0.73和6。
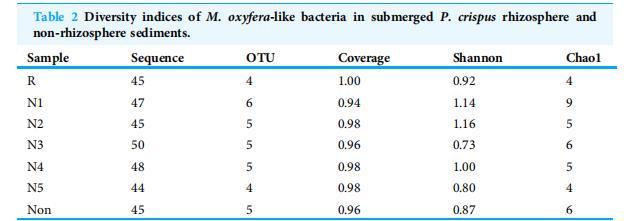
表2 沉水植物菹草根際和非根際沉積物中類產氧甲烷桿菌的多樣性指數。
注:R和Non分別代表根區和非根際區隔室的沉積物樣品;N1-N5代表取自近根際區1至5毫米子隔室的沉積物樣品。
類產氧甲烷桿菌16S rRNA基因的系統發育多樣性
根據Ettwig et al.(2009),OTU1-11的代表性序列被歸類到b群,與產氧甲烷桿菌16S rRNA基因的相似性為88-93%(表3)。b群序列被分類到指定的1至5簇。OTU12代表NC10細菌的一個新簇,不屬于a, b, c或d組(圖2)。
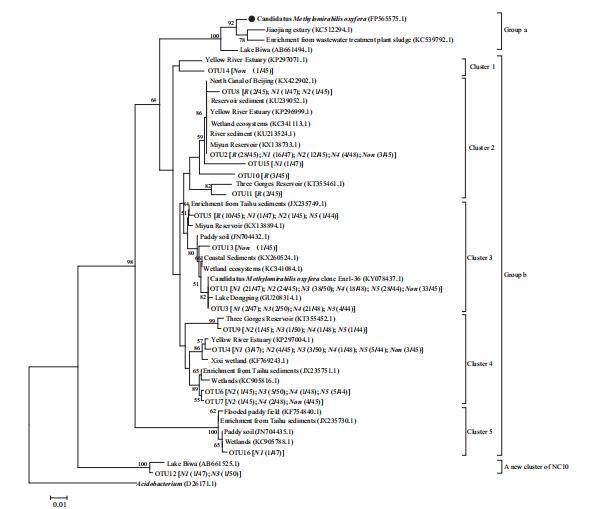
簇1中的序列與非根際隔室的產氧甲烷桿菌16S rRNA基因有92%的相似性。簇2包含OTU2、OTU8、OTU10、OTU11和OTU15。該簇包含來自除N5子隔室(N5)之外所有夾層的73條序列,與產氧甲烷桿菌的相似性為90-92%。簇3是b群中的優勢簇,包含序列數最多(總共324條序列中的205條)。簇3成員在所有夾層均有記錄,與產氧甲烷桿菌的相似性為91-92%。簇4與產氧甲烷桿菌的相似性最高(93%),包含除根區隔室(R)外所有夾層的42條序列。簇5和OTU12在N1和N3近根際子隔室中檢測到,與產氧甲烷桿菌的序列相似性為88-89%(表3)。
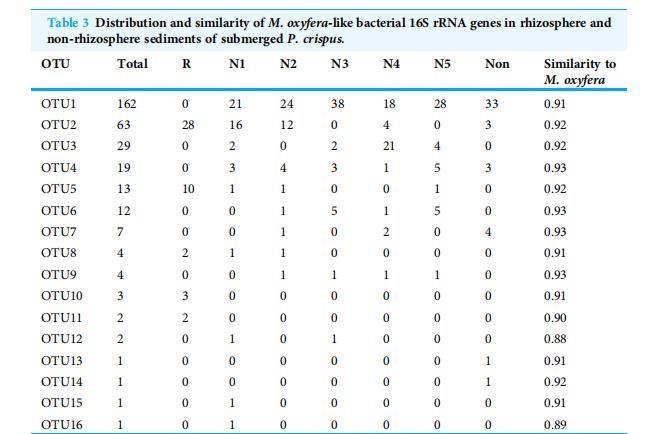
表3 沉水植物菹草根際和非根際沉積物中類產氧甲烷桿菌16S rRNA基因的分布和相似性。
注:R和Non分別代表根區和非根際區隔室的沉積物樣品;N1-N5代表取自近根際區1至5毫米子隔室的沉積物樣品。
討論
在本研究中,我們使用根箱法調查了常見沉水植物根際和非根際區域類產氧甲烷桿菌的出現和多樣性。結果表明,這些細菌存在于根際和非根際沉積物中。大多數OTUs被歸類到b群。每個根箱夾層有4到6個OTUs。細菌群落多樣性指數(香農指數)和豐富度指數(Chao1)分別為0.73-1.16和4-9。
各隔室的多樣性排序為:近根際區 > 根區 > 非根際區(表2),表明近根際區更有利于類產氧甲烷桿菌群落。一些研究發現反硝化細菌(Ruiz-Rueda, Hallin & Ba?eras, 2009),包括厭氧氨氧化細菌(Li et al., 2016)和厭氧甲烷氧化細菌(Vaksmaa et al., 2016)分布在沉水水生植物根系周圍。在我們的研究中,根區和近根際區都具有較高的類產氧甲烷桿菌多樣性,這可能是由于植物根部周圍甲烷和亞硝酸鹽濃度增加所致。植物根系分泌物和植物殘體提供的有機質(OM)可被產甲烷古菌轉化為甲烷(CH?)(Kerdchoechuen, 2005)。此外,根際中的部分反硝化和硝化作用都可能產生亞硝酸鹽(Nie et al., 2015)。這些底物有利于類產氧甲烷桿菌在近根際沉積物中的生長。
本研究七個夾層中獲得的序列與從其他水生生態系統回收的類產氧甲烷桿菌16S rRNA基因序列具有高度相似性。本研究中的簇與河口沉積物(Yan et al., 2015)、濕地(Wang et al., 2012, 2016; Hu et al., 2014)、湖泊(Wang et al., 2015, 2017; Kojima et al., 2012)和厭氧污水污泥(Ma et al., 2017)中的序列密切相關。b群序列主導了菹草根際和非根際區域的類產氧甲烷桿菌(圖2)。本研究未擴增出功能基因pmoA。同樣,在中國東昌湖和東平湖的沉積物中也只發現了類產氧甲烷桿菌16S rRNA基因的b群序列(Wang et al., 2017)。在三峽水庫沉積物中,大多數序列(65/67)屬于b群(Wang et al., 2016),類似地在琵琶湖沉積物(5/6 OTUs屬于b群)(Kojima et al., 2012)和一個城市濕地(8/11 OTUs屬于b群)(Shen et al., 2015)中也觀察到。相反,在椒江口沉積物中,超過87%的類產氧甲烷桿菌16S rRNA基因序列被歸類為a群(Shen et al., 2014b)。沒有證據表明主要由b群細菌組成的群落可以在沒有a群的情況下主導N-DAMO,并且這兩者在大多數自然環境中似乎是共存的。需要進一步研究b群的功能,以增進我們對根際NC10細菌群落的理解。
本研究中類產氧甲烷桿菌的多樣性指數與來自河流(Shen et al., 2014a)和濕地(Wang et al., 2016)的其他淡水沉積物相似。然而,在自然環境中,海洋和河口沉積物中通常發現比淡水沉積物(來自河流和濕地(Yan et al., 2015; Shen et al., 2014a; Wang et al., 2016),特別是湖泊(Deutzmann & Schink, 2011; Kojima et al., 2012; Wang et al., 2017))更高的類產氧甲烷桿菌多樣性。最近,一項關于類產氧甲烷桿菌分布模式的全球分析表明,海洋和淡水生境之間存在顯著的群落差異(Zhang, Liu & Gu, 2018)。因此,群落差異可能是該細菌在海洋中多樣性高于淡水的主要原因。
在本實驗中,同一夾層的樣品被混合后進行測序。雖然混合法能夠檢測到所有的類產氧甲烷桿菌,但該方法無法統計分析不同夾層間OTUs的顯著差異。因此,本文未分析該細菌的空間分布。
結論
在菹草所有三個隔室的沉積物中都檢測到了類產氧甲烷桿菌16S rRNA基因。根際OTU較少,每個夾層僅存在4到6個OTUs。香農指數和Chao1指數與淡水濕地和河流沉積物中的相似。系統發育分析表明,除一個OTU屬于NC10細菌的新簇外,所有OTU均被歸類為類產氧甲烷桿菌16S rRNA基因的b群,這表明b群細菌可能在氮生物地球化學循環中很重要,并可能在調節淡水系統富營養化中發揮重要作用。
 相關新聞
相關新聞