熱線:021-66110810,66110819
手機:13564362870
熱線:021-66110810,66110819
手機:13564362870
3.結果與討論
3.1.pH沖擊對Val-A產量和菌體生長的影響
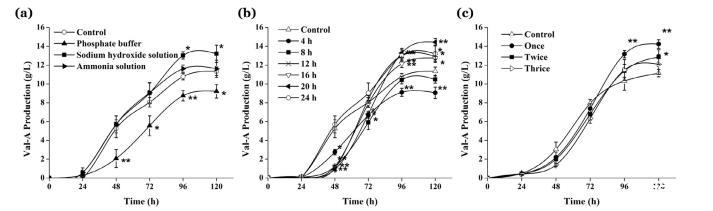
經過初步篩選pH,pH沖擊在8.0時效果最佳。分別向發酵液中添加三種常見的堿性溶液。發現兩種有效的添加劑氫氧化鈉和氨水,而磷酸鹽緩沖液則產生相反的效果(圖1a)。這與一些出版物一致,這些出版物記載過量的無機磷酸鹽通常會抑制次級代謝產物的生產(Sola et al.,2003)。因此,所得結果表明pH沖擊有利于Val-A的生產。氫氧化鈉組的Val-A產量最高,因此選擇氫氧化鈉進行進一步研究。關于pH沖擊的適宜時間,選擇4 h-24 h內的5個時間點作為最旺盛的時期(圖1b)。當堿性沖擊時間晚于24小時時,該處理對Val-A產量沒有顯著影響,未在圖1b中顯示。很明顯,第20小時是進行pH沖擊的最佳時間。在4 h-12 h添加氫氧化鈉時,細胞仍處于生長早期,堿性溶液的刺激在一定程度上阻礙了細胞的生長,因此pH沖擊沒有明顯效果甚至降低了Val-A的產量。至于不同的處理次數,發現多次pH沖擊的效果未達到預期(圖1c)。在Val-A合成過程中,發酵液的pH會持續上升,這使得多次pH沖擊的效果不如單次pH沖擊顯著。總之,最佳條件是在第20小時向發酵液中添加氫氧化鈉溶液,而過多的無機磷酸鹽對Val-A生產有阻礙作用。pH沖擊對吸水鏈霉菌S.hygroscopicus 5008的細胞重量有顯著影響,在96小時時從6.00 g/L增加到6.60 g/L(圖2)。
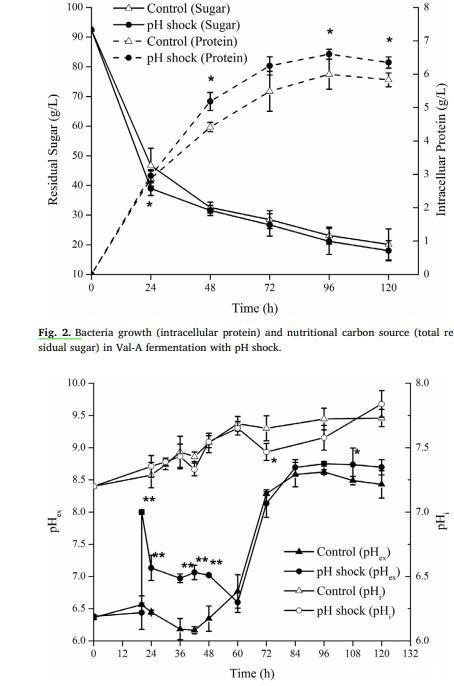
發酵24小時后,pH沖擊組的細胞積累量更高,pH沖擊組的細胞生長速率比對照組快。這一現象表明pH刺激了鏈霉菌的代謝,在一定程度上加速了其生長,同時伴隨著Val-A的大量合成,增加了27.43%。在第四天,pH沖擊組的Val-A產量和產率(13.29 g/L;2.01 g/g蛋白)均高于對照組(10.78 g/L;1.80 g/g蛋白)(圖1c和圖2)。可以推測,在發酵早期積累了前體,然后這些前體被用來合成Val-A。Val-A產量的增加是生物量和產量增加的綜合效應。糖的消耗反映了鏈霉菌的能量代謝。Val-A合成的碳源前體是磷酸戊糖途徑上的景天庚酮糖-7-磷酸(sedoheptulose 7-phosphate)(Degwert et al.,1987;Toyokuni et al.,1987)。pH沖擊組在第一天的糖消耗速率[2.23 g/(L·h)]快于對照組[1.91 g/(L·h)],且pH沖擊組的細胞代謝活性相對較高(圖2)。這主要是由于堿性刺激,細胞加速利用碳水化合物產酸以恢復pH。此外,發酵液中糖的消耗為細胞生長提供了能量,這與生物量的增加相符。
3.2.S.Hygroscopicus 5008培養過程中的pH變化
在5008正常情況下,發酵液pH逐漸升高。在發酵前期,pH緩慢上升,然后快速上升直至達到峰值并趨于穩定(圖3)。發酵液pH是微生物發酵的綜合結果。Val-A可以與氫離子結合,導致發酵液中羥基離子比例上升(Bai et al.,2006)。因此,在發酵后期,當Val-A大量合成時pH上升。在pH沖擊組中,菌絲體首先在自然條件下培養20小時,然后在pH 8.0進行堿性pH沖擊。之后,pH在短時間內迅速下降,這是由脅迫響應引起的。最終,pH在60小時后恢復,進行后續發酵。此外,Val-A在酸性條件下(pH<5.0)可被水解(Jin et al.,2006),因此發酵pH的升高也有利于Val-A的積累和穩定。
胞內pH對微生物的生長和代謝至關重要,它可以影響營養吸收和利用、蛋白質合成、線粒體呼吸、碳水化合物代謝和其他重要的生命活動(Cotter et al.,2001)。因此,只有保持胞內pH的穩定,才能確保微生物的正常生長和代謝。如圖3所示,在整個發酵過程中,胞內pH略有上升,從初始的7.21到最終的7.73。Corvini(Corvini et al.,2004)也使用BCECF-AM熒光探針測定普拉鏈霉菌(Streptomyces pristinaespiralis)的胞內pH,范圍在6.5到8.5之間。還發現基因SHJG7463編碼的陽離子轉運ATP酶在DNA微陣列中高表達,表明陽離子轉運加速以中和pH的變化。此外,大量的ATP將被消耗以轉運細胞膜內外的氫離子,這也解釋了下面關于菌絲體呼吸活性的現象。此外,需要注意的是,三羧酸循環導致外部pH上升,而糖酵解和谷氨酸的合成通常導致酸化。因此,pH沖擊組中的pH波動可能是碳水化合物快速消耗和氨基酸代謝的綜合結果。
3.3.pH沖擊誘導的基因表達全局變化
使用統計標準Fold change大于2且p小于0.05,在菌株5008中鑒定出總共146個pH沖擊下的差異表達基因(DEGs)(表1)。在pH沖擊下顯著變化的DEGs主要分配了代謝、細胞過程和信號傳導以及信息存儲和處理相關的功能。正如預期的那樣,大多數Val-A生物合成基因的轉錄水平在pH沖擊下得到增強,除了葡糖基轉移酶基因valG。
在與代謝相關的DEGs中,基因SHJG7463編碼細胞膜上的陽離子轉運ATP酶。這些基因的上調表達表明鏈霉菌細胞中的轉運蛋白可以及時響應pH沖擊,這有利于營養吸收和次級代謝產物的排泄,并且相關的陽離子轉運蛋白基因與鏈霉菌發酵過程中的pH變化和呼吸鏈密切相關。編碼谷氨酰胺合成酶的SHJG3702上調。這與Gu的研究(Gu et al.,1984;Wei et al.,2012)一致,該研究表明谷氨酰胺和3-氨基-5-羥基苯甲酸(3-Amino-5-hydroxy benzoic acid)可顯著提高Val-A產量,并且谷氨酰胺合成酶活性與Val-A產量呈正相關。另一個DEG SHJG6704編碼銨透性酶(ammonium permease)。夏(Xia and Jiao,1986)發現添加銨離子可以通過加速氮代謝促進Val-A合成過程中的細胞生長。SHJG6704的上調表達為Val-A提供了更多的氮源,并促進了細胞生長。SHJG0557編碼一種細胞色素bd型醌氧化酶(cytochrome bd-type quinol oxidase)。SHJG7814編碼一種糞卟啉原III氧化酶(coproporphyrinogen III oxidase)和相關的Fe-S氧化還原酶。鐵硫氧化還原酶、細胞色素和鐵硫蛋白是細胞電子呼吸鏈的組成部分。SHJG0557和SHJG7814的上調表達意味著吸水鏈霉菌S.hygroscopicus 5008的呼吸發生了變化,這是pH沖擊的響應措施之一。
在與細胞過程和信號轉導相關的DEGs中,編碼UDP-N-乙酰胞壁酸脫氫酶(UDP-N-acetylmuramate dehydrogenase)的基因SHJG3272上調了2.99倍,而編碼細胞分裂蛋白FtsI/青霉素結合蛋白2(cell division protein FtsI/penicillin-binding protein 2)的基因SHJG3894和SHJG3896以及幾個細胞壁水解酶基因則下調。這些基因與細胞壁合成相關,表明在pH沖擊的影響下,細胞壁合成過程得到加強。
在與信息存儲和處理相關的DEGs中,大部分是轉錄調控因子,它們在次級代謝產物合成的調控中發揮作用,也表明S.hygroscopicus 5008通過復雜的信號轉導系統響應pH沖擊。原核生物的基因轉錄需要由核心酶和σ因子組成的RNA聚合酶。編碼RNA聚合酶ECF亞家族σ因子的基因SHJG6022顯著上調。在天藍色鏈霉菌中,雙組分調控系統Cse B/Cse C影響σE(sigma E)的活性(Paget et al.,2002)。環境脅迫信號由雙組分系統傳導。該系統與σE的啟動子結合,然后在核心酶的相互作用下激活轉錄。因此,推測雙組分系統和σE的相互作用也可能在S.hygroscopicus 5008中參與,可以傳遞pH脅迫信號并調控參與Val-A生物合成的基因的表達。
總之,這些DEGs主要與氨基酸代謝、細胞過程和信號傳導以及信息存儲和處理相關。此外,通過qRT-PCR分析驗證了關鍵基因的動態變化。在轉錄變化方面,DNA微陣列和qRT-PCR分析之間顯示出良好的一致性,表明了微陣列數據的可靠性。可以推測,在外源pH沖擊下,細胞壁接收的脅迫信號通過雙組分系統進行轉導。然后由雙組分系統調控的ECFσ因子將與核心酶一起作用于下游基因并激活細胞防御系統。最終,營養物和離子轉運加速,導致葡萄糖代謝、氨基酸代謝、呼吸鏈和其他生理活動的變化,同時也緩解了堿性刺激并促進了Val-A的生物合成。

表1 pH沖擊后5008中的差異表達基因:使用COG進行分類
3.4.Val-A發酵過程中呼吸活性的變化
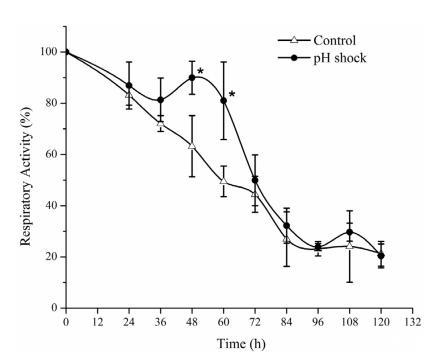
如圖4所示,兩種策略下的菌絲體呼吸活性在接種后均下降,這可能歸因于程序性細胞死亡。36小時后,由于蛋白質的快速合成和更多的細胞積累,pH沖擊組的呼吸活性持續增加了12小時,然后開始下降。微生物呼吸鏈成分的基礎是NAD、FAD、FAM、鐵硫蛋白(Fe-S)、泛醌(輔酶Q)和細胞色素系統。上調表達的基因SHJG0557編碼細胞色素BD氧化酶,SHJG7814編碼鐵硫氧化還原酶,這些酶是呼吸鏈的基本組成部分。可以推測,pH沖擊后呼吸電子傳遞鏈中的酶活性增強,因此電子傳遞和呼吸加速以產生更多能量,這對于Val-A合成至關重要。
3.5.pH沖擊對氮代謝的影響
先前的研究使用氮同位素標記法報道,谷氨酸很可能是氮橋的主要來源(Mahmud et al.,1999)。根據Hodgson的報告(Hodgson,2000)和KEGG,S.hygroscopicus 5008的氮同化途徑如圖5所示。當氮源充足時,銨被谷氨酸脫氫酶催化成谷氨酸。當缺乏氮源時,銨主要被谷氨酰胺合成酶(GS)催化成谷氨酰胺,然后谷氨酰胺轉化為谷氨酸。谷氨酰胺和谷氨酸都參與了Val-A生物合成的一系列反應過程。微陣列和qRT-PCR的結果顯示,基因SHJG3702和SHJG6704均上調。隨后,我們發現pH沖擊組胞內谷氨酸濃度是對照組的2.82倍(圖5),這表明在pH沖擊下銨離子轉運加速,從而產生更多的谷氨酰胺和谷氨酸。顯然,細胞處于相對高氮的環境中,而Val-A合成正在進行,因此更多的氮前體(谷氨酸)有利于Val-A的合成。此外,先前對一系列微生物的研究表明,輸入的肽可以被分解成游離氨基酸,形成重要的質子來源(Giotis et al.,2010)。谷氨酸是酸性氨基酸,增加的谷氨酸除了提供氮源外,還可以緩解堿的壓力。
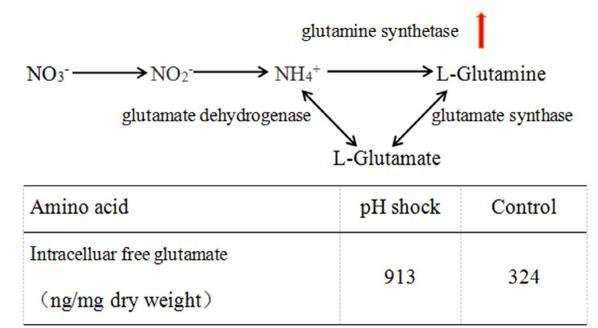
圖5.pH沖擊后5008的氮同化途徑和胞內游離谷氨酸的變化。
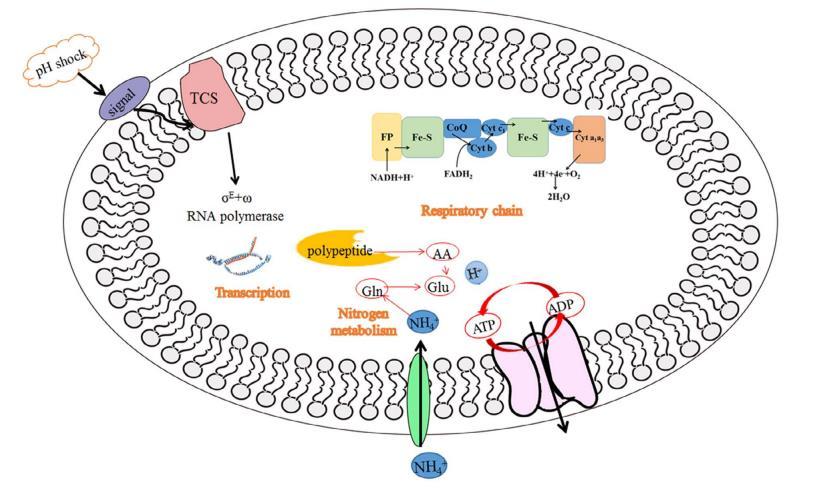
4.結論
堿性pH沖擊使Val-A產量顯著提高了27.43%。結合DNA微陣列,研究了其對細胞呼吸活性和胞內微環境的影響,揭示了鏈霉菌對pH沖擊的細胞響應機制(圖6)。pH沖擊加速了質子轉運并增強了細胞的呼吸活性,從而為細胞生長提供了更多能量。并且此過程中酸性氨基酸的形成為井岡霉素合成提供了更多的氮前體。因此,上述因素的共同作用提高了Val-A的產量。顯然,在鏈霉菌發酵中應更加重視堿性pH沖擊。
致謝
本工作得到浙江省公益項目(2016C32007)、國家重點研發計劃(2016YFD0401201)、浙江省科協青年人才培育項目(2016YCGC004)和國家自然科學基金(20906060)的資助。
 相關新聞
相關新聞