熱線:021-66110810,66110819
手機(jī):13564362870
熱線:021-66110810,66110819
手機(jī):13564362870
摘要缺氧(hypoxia)是細(xì)菌在宿主定植過程中面臨的常見挑戰(zhàn),部分原因是形成了密集聚集的群落(生物膜)。cbb3型細(xì)胞色素c氧化酶,催化呼吸作用的終末步驟并具有高氧親和力,已被認(rèn)為與細(xì)菌致病性相關(guān)。假單胞菌屬不同尋常之處在于它們通常包含多個(gè)完整和部分(即“孤兒”)的cbb3型氧化酶和氧化酶亞基操縱子。在此,我們描述了孤兒催化亞基CcoN4在機(jī)會(huì)性病原體銅綠假單胞菌PA14菌株的菌落生物膜發(fā)育和呼吸作用中的獨(dú)特作用。我們還發(fā)現(xiàn),CcoN4有助于還原吩嗪(phenazines)——一種在生物膜中支持細(xì)胞氧化還原平衡的抗生素,并有助于在秀麗隱桿線蟲(Caenorhabditis elegans)感染模型中的毒力。這些結(jié)果突顯了菌落生物膜模型與致病性的相關(guān)性,并強(qiáng)調(diào)了cbb3型氧化酶作為治療靶點(diǎn)的潛力。
引言
在可用于生物還原的氧化劑中,分子氧(O2)提供最高的自由能產(chǎn)量。自從O2在大約24億年至5.4億年前在大氣中積累以來(Kirschvink and Kopp,2008;Dietrich et al.,2006b),能夠利用它生長(zhǎng)和生存并耐受其有害副產(chǎn)物的生物已經(jīng)進(jìn)化以利用這種能量并增加了復(fù)雜性(Knoll and Sperling,2014;Falkowski,2006)。在小尺度和擁擠的環(huán)境中,O2的快速消耗導(dǎo)致對(duì)該資源的競(jìng)爭(zhēng),并促進(jìn)了細(xì)菌和古細(xì)菌的O2還原機(jī)制多樣化,而這種多樣化在真核生物中沒有發(fā)生(Brochier-Armanet et al.,2009)。使細(xì)菌能夠呼吸O2的各種酶表現(xiàn)出不同的親和力和質(zhì)子泵效率,并可能有助于在缺氧生態(tài)位中的競(jìng)爭(zhēng)成功(Morris and Schmidt,2013)。此類環(huán)境包括被具有高度農(nóng)業(yè)(Preisig et al.,1996)和臨床(Way et al.,1999;Weingarten et al.,2008)重要性的細(xì)菌定植的動(dòng)植物宿主組織。
機(jī)會(huì)性病原體銅綠假單胞菌(Pseudomonas aeruginosa),一種能夠定植于植物和動(dòng)物宿主的細(xì)菌(Rahme et al.,1995),具有分支的呼吸鏈,具有使用五種經(jīng)典的末端氧化酶復(fù)合物將O2還原為水的潛力:兩種醌氧化酶(bo3(Cyo)和一種氰化物不敏感bd型氧化酶(CIO))和三種細(xì)胞色素c氧化酶(aa3(Cox)、cbb3-1(Cco1)和cbb3-2(Cco2))(圖1A)。幾篇關(guān)鍵出版物描述了銅綠假單胞菌的末端氧化酶補(bǔ)體,揭示了該生物特有的特征(Williams et al.,2007;Comolli and Donohue,2004;Alvarez-Ortega and Harwood,2007;Arai et al.,2014;Kawakami et al.,2010;Jo et al.,2014)。銅綠假單胞菌的不同尋常之處在于它編碼兩種屬于cbb3型家族的氧化酶。這些酶以其在低O2濃度下相對(duì)較高的催化活性以及僅限于細(xì)菌域(Brochier-Armanet et al.,2009;Pitcher and Watmough,2004)而著稱。(銅綠假單胞菌的cbb3型氧化酶通常被稱為cbb3-1和cbb3-2;然而,我們將使用‘Cco1’和‘Cco2’來表示這些酶,與其編碼基因的注釋一致。)大多數(shù)編碼cbb3型氧化酶的細(xì)菌基因組只包含一個(gè)此類復(fù)合物的操縱子,其在O2限制條件下特異性誘導(dǎo)(Cosseau and Batut,2004)。在銅綠假單胞菌中,cco2操縱子在低O2濃度生長(zhǎng)期間被誘導(dǎo),但cco1操縱子在高水平組成型表達(dá)(Comolli and Dono-hue,2004;Kawakami et al.,2010)。
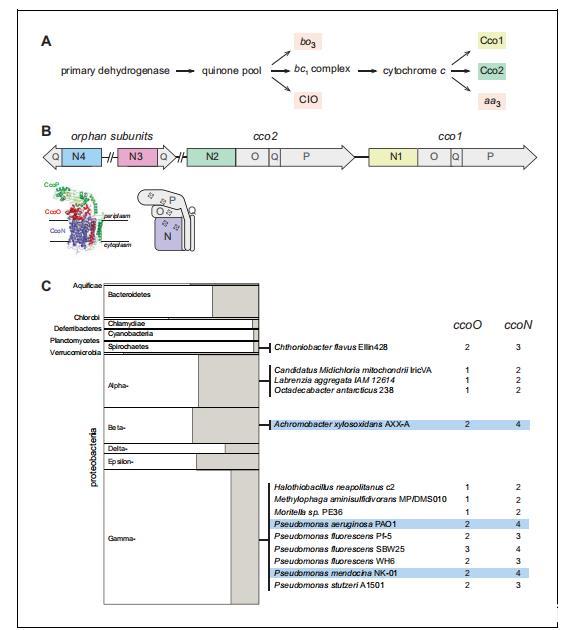
圖1銅綠假單胞菌的呼吸鏈和cco基因及蛋白產(chǎn)物的排列,以及ccoN孤兒基因的系統(tǒng)發(fā)育分布。(A)銅綠假單胞菌的分支電子傳遞鏈,包含五個(gè)典型的末端氧化酶。(B)cco基因在銅綠假單胞菌基因組中的組織結(jié)構(gòu)。Cco復(fù)合物的圖解基于P.stutzeri的Cco結(jié)構(gòu)(PDB:3mk7)。(C)左圖:各細(xì)菌門中含有ccoO和N同源物的基因組比例示意圖。由于Chrysiogenetes、Gemmatimonadetes和Zetaproteobacteria支系中只有一個(gè)物種含有ccoO和N同源物,因此省略了這些支系。
銅綠假單胞菌末端氧化酶補(bǔ)體的另一個(gè)奇特之處在于,在染色體上遠(yuǎn)離cco1和cco2操縱子的位置存在編碼“孤兒”cbb3型亞基的基因。雖然染色體相鄰的cco1和cco2操縱子各自包含四個(gè)編碼功能性Cco復(fù)合物(由N、O、P和Q亞基組成)的基因,但另外兩個(gè)部分操縱子ccoN3Q3和ccoN4Q4各自僅包含編碼Q和催化性N亞基的同源基因(圖1B)。ccoN3Q3操縱子的表達(dá)在厭氧反硝化條件下被誘導(dǎo)(Alvarez-Ortega and Harwood,2007),并在2%O2下生長(zhǎng)期間暴露于亞硝酸鹽時(shí)被誘導(dǎo)(Hirai et al.,2016)。在液體培養(yǎng)物中需氧生長(zhǎng)期間,ccoN4Q4被氰化物誘導(dǎo),氰化物在穩(wěn)定期產(chǎn)生(Hirai et al.,2016)。然而,額外的表達(dá)研究表明ccoN4Q4轉(zhuǎn)錄受氧化還原條件影響,因?yàn)樵摬倏v子被O2限制誘導(dǎo)(Alvarez-Ortega and Harwood,2007)并在響應(yīng)銅綠假單胞菌產(chǎn)生的氧化還原活性抗生素pyocyanin(綠膿菌素)時(shí)輕微下調(diào)(Dietrich et al.,2006a)。
在最近的一項(xiàng)研究中,Hirai等人表征了銅綠假單胞菌PAO1中含有經(jīng)典亞基和孤兒亞基組合的cbb3異構(gòu)體的生化特性和生理作用(Hirai et al.,2016)。在一個(gè)缺乏所有需氧末端氧化酶的菌株中,表達(dá)任何異構(gòu)體都賦予利用O2生長(zhǎng)的能力,證實(shí)含有孤兒N亞基的異構(gòu)體具有功能。此外,作者發(fā)現(xiàn),在低O2條件下的液體培養(yǎng)物生長(zhǎng)期間,ccoN3Q3和ccoN4Q4的產(chǎn)物分別有助于對(duì)亞硝酸鹽和氰化物的抗性。雖然這些結(jié)果為cbb3異質(zhì)復(fù)合物在液體培養(yǎng)物生長(zhǎng)中的作用提供了見解,但含N3和N4的異構(gòu)體在生物膜生長(zhǎng)和致病性中的潛在作用仍有待探索。
生物膜生活方式——細(xì)胞在自產(chǎn)基質(zhì)包裹的密集群落中生長(zhǎng)——已被認(rèn)為與多種系統(tǒng)中感染的確立和持續(xù)存在有關(guān)(Edwards and Kjellerup,2012;Rybtke et al.,2015)。生物膜發(fā)育促進(jìn)了O2梯度的形成,使得遠(yuǎn)離生物膜表面的細(xì)胞處于缺氧或無氧條件(Werner et al.,2004)。使用菌落形態(tài)分析來研究氧化還原代謝及其與群落行為的關(guān)系,我們發(fā)現(xiàn)生物膜中細(xì)胞的O2限制導(dǎo)致細(xì)胞內(nèi)氧化還原狀態(tài)失衡。這可以通過群落形態(tài)的改變來緩解,該改變?cè)黾恿松锬さ谋砻娣e與體積比,從而增加了常駐細(xì)胞對(duì)O2的獲取(Kempes et al.,2014)。對(duì)于生物膜中的銅綠假單胞菌細(xì)胞,還原力的細(xì)胞內(nèi)積累也可以通過產(chǎn)生和還原內(nèi)源性抗生素吩嗪來防止,吩嗪介導(dǎo)向遠(yuǎn)處可用的氧化劑的胞外電子轉(zhuǎn)移(Dietrich et al.,2013)。我們發(fā)現(xiàn)生物膜特異性的吩嗪產(chǎn)生有助于急性肺感染小鼠模型中的致病性(Recinos et al.,2012),進(jìn)一步說明了吩嗪介導(dǎo)的氧化還原平衡對(duì)于群落中銅綠假單胞菌細(xì)胞的重要性。
由于生物膜生活方式固有的O2梯度形成,我們假設(shè)銅綠假單胞菌cco操縱子的差異調(diào)節(jié)影響了它們對(duì)生物膜亞區(qū)代謝電子流的貢獻(xiàn)。我們?cè)u(píng)估了各種cbb3型氧化酶異構(gòu)體在多細(xì)胞行為和毒力中的作用。我們的結(jié)果表明,含有孤兒亞基CcoN4的異構(gòu)體可以通過O2和吩嗪還原支持生物膜中的生存,并有助于銅綠假單胞菌在秀麗隱桿線蟲“緩慢致死”感染模型中的致病性。
 相關(guān)新聞
相關(guān)新聞