熱線:021-66110810,66110819
手機:13564362870
熱線:021-66110810,66110819
手機:13564362870
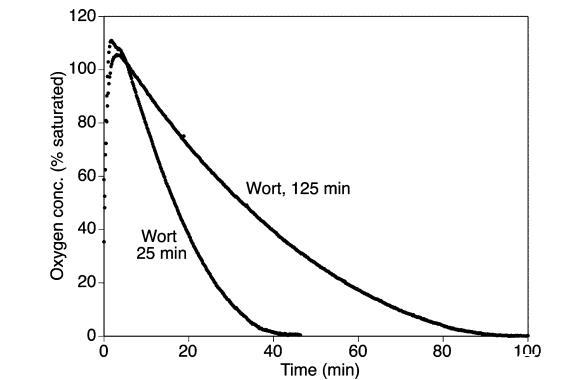
通過測量麥汁樣品在加熱到60°C時消耗溶解氧的能力,也檢查了麥汁樣品的氧化穩定性(圖5)。早期麥汁樣品消耗氧氣的速率高于糖化結束時收集的麥汁。這與基于ESR的自旋捕獲方法觀察到的相對氧化穩定性一致,因此證明添加乙醇和自旋捕獲劑POBN不會引起麥汁樣品氧化行為的重大變化。麥汁樣品在60°C時消耗氧氣很快。在耗氧量測量前預熱樣品會導致測量開始時樣品的氧含量水平差異很大。因此,最可重復的結果是在將室溫麥汁樣品加入預熱的氧測量室時開始測量獲得的。然而,這導致了在樣品溫度達到60°C的前10分鐘測量期間觀察到的氧氣濃度迅速變化(圖5)。
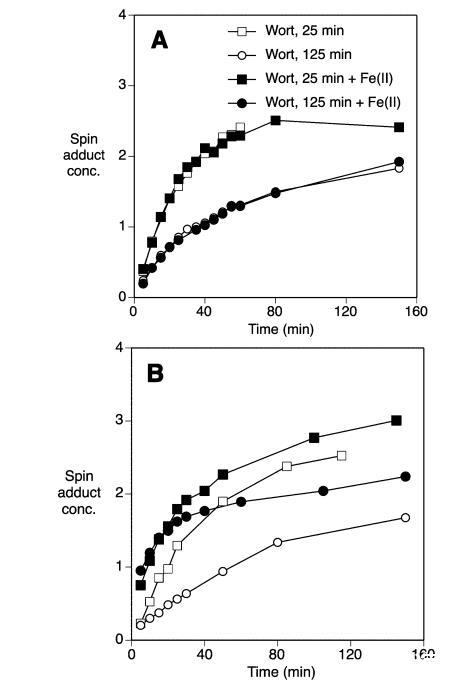
還測試了早期麥汁樣品在培養過程中較高的自由基形成速率是否是由于存在內源麥芽氧化還原活性酶。脂氧合酶(LOX-1和-2)在高于65°C的溫度下失活,預計在應用的糖化程序的最后一步中失活。在評估氧化穩定性期間,在70°C下培養麥汁樣品,所有樣品的自由基形成速率大約提高了四倍。然而,在兩種培養溫度下,來自糖化早期低溫階段的麥汁樣品都比后期高溫糖化階段的樣品顯示出更高的自由基形成速率(圖4)。這表明早期樣品較低的氧化穩定性很可能不是由麥芽氧化還原活性酶的作用引起的
在麥汁樣品培養期間的作用。然而,有可能在糖化的早期低溫階段,氧化還原酶產生了在麥汁加熱時產生自由基的前體。在這種情況下,這些潛在的前體可能在糖化的最后高溫階段被破壞,從而產生在ESR評估自由基形成期間加熱時不易產生自由基的后期麥汁樣品。
糖化過程中添加鐵:對麥汁樣品中鐵和銅水平的分析表明,最終麥汁比糖化25分鐘后收集的麥汁含有更少的鐵和銅(表1)。過渡金屬,特別是鐵和銅的存在,可以促進金屬催化的氧化,從而降低氧化穩定性。在醪液開始時添加鐵并不影響麥汁樣品中自旋加合物的形成(圖6),但在糖化和過濾后直接將鐵添加到麥汁中顯著增加了自旋加合物的水平。
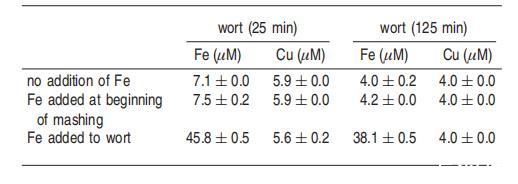
麥汁樣品中的鐵濃度分析表明,在糖化過程中添加鐵僅略微增加了麥汁中的鐵水平,而直接向麥汁樣品中添加金屬則顯著提高了鐵濃度(表1)。這些結果表明鐵在糖化過程中被有效結合。
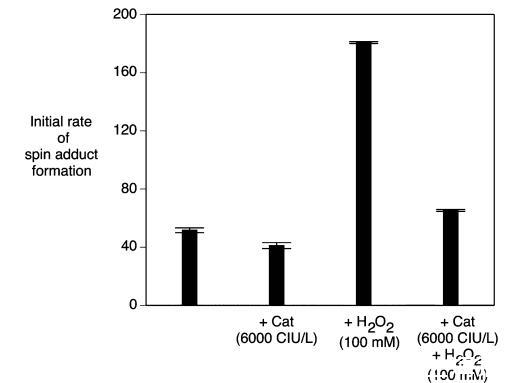
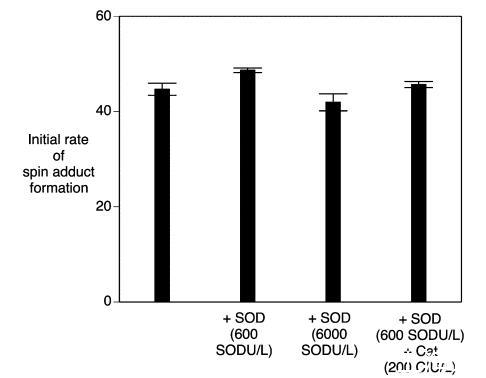
過氧化氫酶和超氧化物歧化酶:過氧化氫和超氧化物是啤酒氧化過程中重要的中間體。過氧化氫酶能有效去除過氧化氫,并且已證明向啤酒中添加過氧化氫酶可抑制自由基的形成。向麥汁樣品中添加過氧化氫酶(6000 CIU/L)導致自由基形成速率顯著降低了20%(p<0.05)(圖7)。正如預期的那樣,添加過氧化氫顯著增加了自由基的形成(圖7)。同時添加過氧化氫和過氧化氫酶降低了自旋加合物的形成速率,證明過氧化氫酶在麥汁樣品培養使用的高溫(60°C)下是有效的。同樣檢查了超氧化物歧化酶的效果。盡管如此,單獨添加超氧化物歧化酶或與過氧化氫酶聯合添加,與對照相比,對自由基形成速率均未產生顯著的正向或負向影響(p>0.05)(圖8)。當超氧化物歧化酶與顯著較低劑量的過氧化氫酶(200 CIU/L)聯合使用時缺乏效果可能是原因。在低過氧化氫底物濃度下,如果存在合適的受體(如有機底物),過氧化氫酶能夠充當過氧化物酶。這是由于過氧化氫酶和過氧化物酶對過氧化氫的親和力存在巨大差異。過氧化氫酶在低底物濃度下充當過氧化物酶的可能性或許可以解釋為什么
在明顯較低劑量范圍內沒有觀察到過氧化氫酶的效果。
討論
通過測量甜麥汁樣品在有空氣接觸加熱過程中自由基形成的傾向來評估其氧化穩定性。自由基通過自旋捕獲技術檢測。然而,乙醇的存在對于積累濃度足以產生足夠強度ESR信號的自旋加合物是必要的。乙醇對麥汁中自旋加合物形成的影響表明,在麥汁加熱過程中形成了高活性自由基,如羥基自由基和烷氧基自由基。這些自由基能夠與乙醇反應生成1-羥乙基自由基,后者被POBN捕獲(反應1-3)。
因此,在自旋捕獲實驗中使用乙醇不僅增加了ESR可檢測的自旋加合物的形成,而且使得對高活性自由基的檢測更具選擇性。此外,加合物{}^{bullet}POBN/CHleft(CH_{3}right)OH良好的熱穩定性允許在麥汁高溫培養過程中積累相對高濃度的該自旋加合物,從而產生高強度的ESR譜圖。在無乙醇情況下觀察到的ESR譜低強度不太可能是由于麥汁中缺乏自由基形成。在沒有乙醇的情況下,非選擇性的羥基和烷氧基自由基僅與麥汁中存在的碳水化合物、蛋白質、多酚和其他有機成分反應,但這些化合物衍生的自由基顯然不形成自旋加合物,或者形成的自旋加合物不如乙醇衍生的加合物穩定。羥基自由基與碳水化合物的反應通常不具有選擇性,可能導致多種產物,包括解聚和開環產物。
已證明氧是自由基形成所必需的。酶促反應,如脂氧合酶活性,在本研究麥汁加熱過程中發生的氧化過程中很可能作用可忽略,因為它們在高于65°C的溫度下失活。生物系統中羥基和烷氧基自由基的產生通常與鐵和銅離子的存在密切相關,這些離子能夠通過芬頓反應(反應4)將過氧化物轉化為這些自由基。
有機過氧化物ROOH(如蛋白質過氧化物)可能通過類似反應與Fe(II)反應,產生烷氧基自由基(反應5)。
相應地,向麥汁樣品中添加Fe(II)確實增加了自旋加合物的形成。值得注意的是,在糖化開始時添加鐵并沒有增加麥汁樣品中的鐵含量。顯然,麥汁中的高鐵含量通過其與固形物的有效結合而被阻止檢測到,這些固形物在糖化室的過濾或壓濾過程中被去除。
羥基和烷氧基自由基的形成表明過氧化物作為前體存在于麥汁中。因此,向麥汁中添加過氧化氫增加了自由基的形成,而過氧化氫酶(即使在麥汁培養使用的高溫下也能非常有效地去除過氧化氫)能夠阻止由添加過氧化氫引起的額外自由基形成。然而,在麥汁培養期間單獨添加過氧化氫酶對麥汁中自由基的形成速率有顯著影響,將其降低了20%。這表明以過氧化氫為試劑的芬頓反應(反應4)并非麥汁中氧化事件唯一的反應機制。此外,添加SOD對麥汁中的自由基形成沒有正向或負向影響,這表明超氧化物在有氧氧化過程中不是重要的中間體。
兩種機制可能解釋自由基的形成無需游離過氧化氫或超氧化物作為中間體。有人提出,高活性氧酰離子(ferryl ions)Fe^{IV}=O可以通過氧作為雙鐵-氧絡合物結合(反應6-8)的機制產生。已經表明,過氧化氫酶和SOD僅能部分防止鐵自氧化過程中氧化物種的形成。這些物種中的一些,非常可能包括氧酰離子Fe^{IV}=O,取決于存在的螯合劑,能夠將乙醇氧化成1-羥乙基自由基,后者已被檢測為自旋加合物。
可以提出另一種機制,其中源自有機化合物(如碳水化合物或蛋白質)的烷基自由基捕獲氧氣形成過氧自由基(反應9)。麥汁中還原性化合物(如多酚)的存在導致過氧自由基快速轉化為氫過氧化物(反應10),后者是形成烷氧基自由基的前體(反應5)。烷氧基自由基隨后與碳水化合物或蛋白質反應,形成新的烷基自由基(反應11)。
根據目前的結果,無法區分這些機制。然而,這兩種機制很可能并行發生,其中通過反應6-8產生的氧酰離子與碳水化合物或蛋白質反應,形成捕獲氧氣(反應9)的烷基自由基。
麥汁樣品加熱后立即形成自由基,這與貯藏啤酒(lager beer)形成對比,后者通常觀察到自由基形成前有一個滯后期。貯藏啤酒風味穩定性與這些自由基形成滯后期長度之間存在良好的相關性。滯后期被認為是由于促氧化成分和抗氧化劑(如亞硫酸鹽)的競爭效應所致。啤酒中的亞硫酸鹽可以有效淬滅過氧化物,而過氧化物是自由基形成的重要前體。麥汁中自由基形成缺乏滯后期,表明麥汁中的內源抗氧化劑僅能淬滅一部分自由基,留下大量自由基參與有害的氧化反應。在麥汁過濾(mashing-off)時向麥汁中添加亞硫酸鹽未觀察到自由基形成的滯后期,表明亞硫酸鹽在麥汁中會快速衰減。
在本研究中,使用甜麥汁中自旋加合物形成的初始速率來量化和研究糖化過程中氧化反應的發生程度。然而,需要進一步研究來評估啤酒釀造早期步驟中氧化的影響如何受到后續釀造步驟的調節,最重要的是,它們如何影響最終啤酒產品的質量。這包括進一步研究過氧化氫酶促進的甜麥汁自旋加合物初始形成速率降低對最終啤酒風味穩定性的影響。
 相關新聞
相關新聞