熱線:021-66110810,66110819
手機(jī):13564362870
熱線:021-66110810,66110819
手機(jī):13564362870
Modification of oxygen consumption and blood flow in mouse somatosensory cortex by cell-type-specific neuronal activity
細(xì)胞類(lèi)型特異性神經(jīng)元活動(dòng)對(duì)小鼠體感皮層耗氧量和血流的改變
來(lái)源:Journal of Cerebral Blood Flow & Metabolism 2020, Vol. 40(10) 2010–2025
1. 論文摘要核心內(nèi)容
本研究通過(guò)光遺傳學(xué)技術(shù)特異性激活皮層PV中間神經(jīng)元和錐體神經(jīng)元,探究γ振蕩(gamma activity)與腦氧代謝率(CMRO?)的關(guān)系:
核心發(fā)現(xiàn):CMRO?反應(yīng)取決于神經(jīng)元激活本身,而非γ活動(dòng)功率(圖6)。光遺傳激活PV中間神經(jīng)元可獨(dú)立誘導(dǎo)CMRO?和腦血流(CBF)反應(yīng),但γ活動(dòng)本身并非能量消耗的直接驅(qū)動(dòng)因素。
關(guān)鍵機(jī)制:
錐體神經(jīng)元激活是γ振蕩產(chǎn)生的必要條件(圖3, 5);
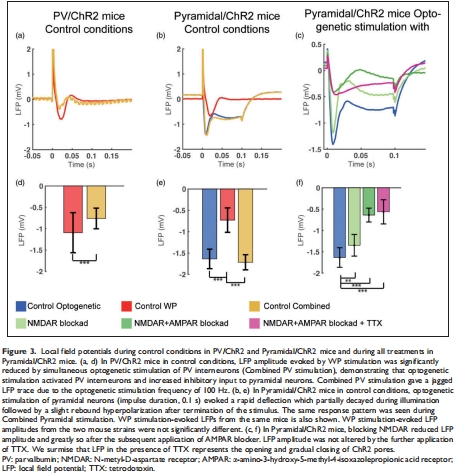
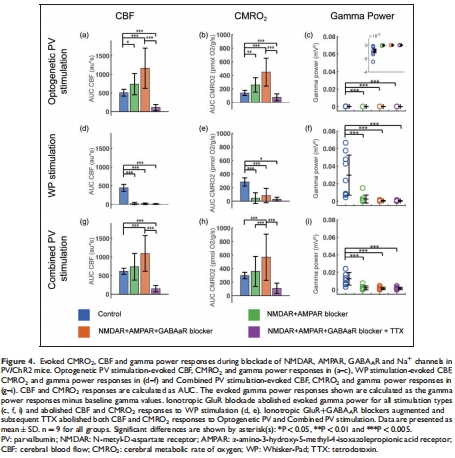
共興奮-抑制(如PV中間神經(jīng)元抑制錐體神經(jīng)元)不增強(qiáng)能量消耗(圖4, 7);
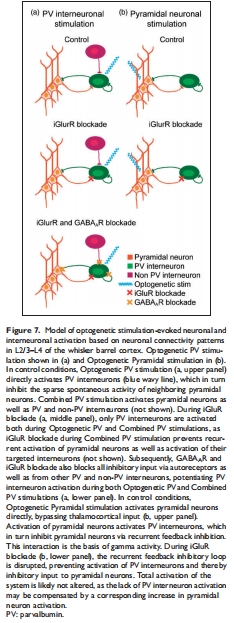
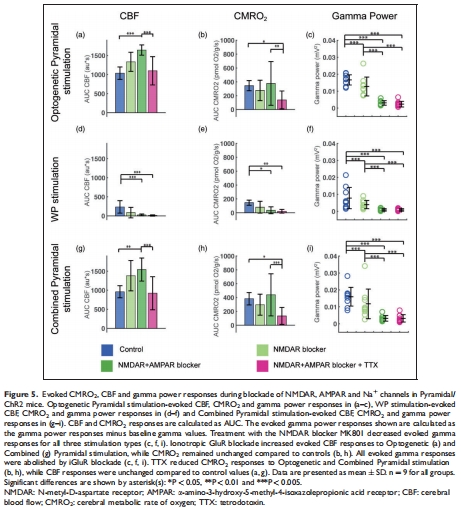
CMRO?主要反映電壓門(mén)控鈉通道活動(dòng)(圖4, 5)。
創(chuàng)新點(diǎn):首次在細(xì)胞類(lèi)型分辨率下證明神經(jīng)元興奮而非節(jié)律活動(dòng)是腦能量消耗的核心驅(qū)動(dòng)因素。
2. 研究目的
1.驗(yàn)證假說(shuō):γ振蕩(30-90 Hz)是否直接驅(qū)動(dòng)高能量消耗?PV中間神經(jīng)元激活是否通過(guò)共興奮-抑制增強(qiáng)氧耗?
2.解析機(jī)制:明確PV中間神經(jīng)元在神經(jīng)血管耦合(CBF-CMRO?匹配)中的作用。
3.技術(shù)突破:結(jié)合光遺傳學(xué)與丹麥Unisense微電極,實(shí)現(xiàn)細(xì)胞類(lèi)型特異性的CMRO?動(dòng)態(tài)監(jiān)測(cè)。
3. 研究思路
1.動(dòng)物模型:
轉(zhuǎn)基因小鼠:PV-ChR2(PV中間神經(jīng)元特異性表達(dá)光敏感通道)、Pyramidal-ChR2(錐體神經(jīng)元特異性表達(dá))。
刺激方式:光遺傳激活PV/錐體神經(jīng)元、觸須墊刺激(Whisker-Pad, WP)、聯(lián)合刺激(圖1c-g)。
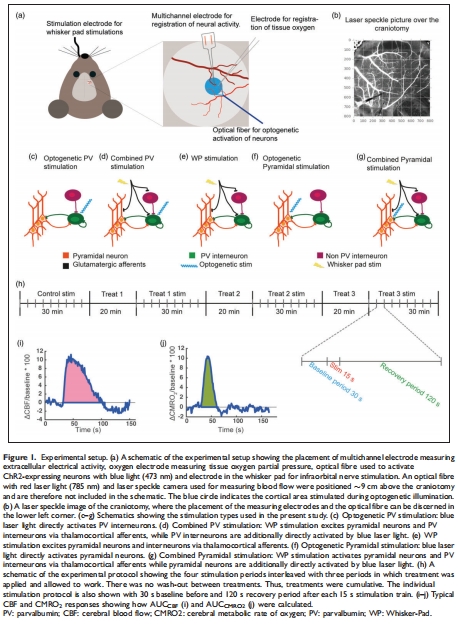
2.干預(yù)手段:
藥理學(xué)阻斷:NMDAR/AMPAR(谷氨酸受體)、GABAAR、電壓門(mén)控鈉通道(TTX)。
3.多模態(tài)監(jiān)測(cè):
丹麥Unisense電極:實(shí)時(shí)測(cè)量局部組織氧分壓(tpO?),計(jì)算CMRO?(方法2.6);
激光散斑成像(LSCI):腦血流(CBF)動(dòng)態(tài);
電生理:局部場(chǎng)電位(LFP)及γ功率分析。
4. 關(guān)鍵數(shù)據(jù)及研究意義
(1) PV中間神經(jīng)元激活獨(dú)立驅(qū)動(dòng)CMRO?(圖2, 4)
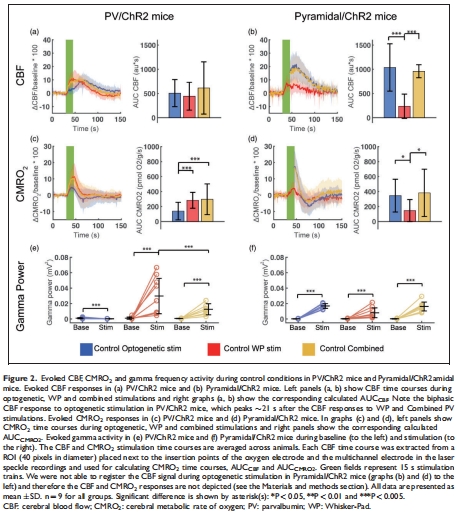
數(shù)據(jù):
光遺傳激活PV神經(jīng)元誘導(dǎo)顯著CMRO?上升(ΔCMRO? ≈ 40%,圖2c);
iGluR/GABAAR阻斷后CMRO?反應(yīng)仍存在,TTX阻斷后消失(圖4a-b),證明其依賴電壓門(mén)控鈉通道。
意義:PV神經(jīng)元自身活動(dòng)即可消耗氧氣,無(wú)需錐體神經(jīng)元參與,推翻“抑制性神經(jīng)元僅通過(guò)抑制消耗能量”的傳統(tǒng)認(rèn)知。
(2) γ振蕩與CMRO?解離(圖6)
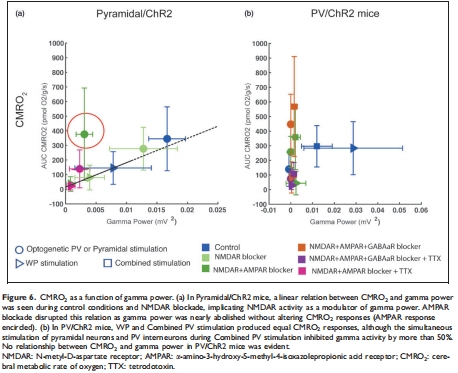
數(shù)據(jù):
錐體神經(jīng)元激活時(shí),γ功率與CMRO?呈正相關(guān)(圖6a);
但AMPAR阻斷后γ功率↓80%而CMRO?不變(圖6a紅圈);PV激活時(shí)γ↓50%但CMRO?不變(圖6b)。
意義:γ活動(dòng)是能量消耗的伴隨現(xiàn)象而非原因,顛覆“γ振蕩是高能耗過(guò)程”的假說(shuō)。
(3) 共興奮-抑制不增強(qiáng)能量消耗(圖4g-h, 7)
數(shù)據(jù):聯(lián)合刺激(WP+PV光遺傳)抑制錐體神經(jīng)元活動(dòng)(LFP↓50%,圖3a),但CMRO?與單純WP刺激無(wú)差異(圖4g-h)。
意義:反駁“興奮-抑制平衡增強(qiáng)跨膜離子通量”的理論(圖7模型),表明抑制性輸入不額外增加氧耗。
(4) 神經(jīng)血管解耦現(xiàn)象(圖5)
數(shù)據(jù):
iGluR阻斷后,錐體神經(jīng)元光遺傳刺激的CBF↑但CMRO?不變(圖5a-b);
TTX阻斷后CMRO?↓但CBF不變(圖5h)。
意義:CBF與CMRO?可短暫解耦,提示血流反應(yīng)可能受非代謝因素(如神經(jīng)遞質(zhì))調(diào)控。
5. 丹麥Unisense電極的核心價(jià)值
(1) 技術(shù)原理
功能:高時(shí)空分辨率測(cè)量皮層氧分壓(tpO?)(方法2.6),結(jié)合CMRO?計(jì)算模型量化腦氧代謝率。
參數(shù):
尖端直徑10 μm,空間分辨率20 μm(精準(zhǔn)定位L2/3層);
采樣率100 Hz → 1 Hz(降噪保真)。
(2) 關(guān)鍵發(fā)現(xiàn)(圖2c-d, 4, 5b)
細(xì)胞類(lèi)型特異性CMRO?動(dòng)態(tài):
PV神經(jīng)元激活后tpO?快速下降(圖2c),直接證明其獨(dú)立耗氧能力;
錐體神經(jīng)元激活后CMRO?反應(yīng)依賴鈉通道(TTX阻斷后↓66%,圖5b)。
γ振蕩與氧耗解耦的直接證據(jù):
AMPAR阻斷后γ功率消失但tpO?動(dòng)態(tài)不變(圖5b vs 圖6a),為“γ非耗能主因”提供原位證據(jù)。
(3) 研究意義
機(jī)制解碼:
推翻“γ振蕩驅(qū)動(dòng)高能耗”假說(shuō),確立神經(jīng)元興奮為氧耗核心;
揭示PV神經(jīng)元自身代謝需求,修正抑制性神經(jīng)元功能模型。
技術(shù)優(yōu)勢(shì):
精準(zhǔn)性:微米級(jí)空間分辨率避免組織平均效應(yīng);
動(dòng)態(tài)性:秒級(jí)監(jiān)測(cè)捕捉瞬態(tài)氧代謝變化;
不可替代性:傳統(tǒng)fMRI/BOLD無(wú)法區(qū)分細(xì)胞類(lèi)型特異性代謝。
6. 結(jié)論
1.γ振蕩本質(zhì):神經(jīng)元興奮的副產(chǎn)品,非獨(dú)立耗能過(guò)程。
2.PV神經(jīng)元作用:自身活動(dòng)直接耗氧,抑制錐體神經(jīng)元不額外增加能量需求。
3.神經(jīng)血管耦合:CBF與CMRO?可解耦,血流反應(yīng)或受非代謝信號(hào)調(diào)控。
4.Unisense電極貢獻(xiàn):
實(shí)現(xiàn)細(xì)胞類(lèi)型分辨的CMRO?動(dòng)態(tài)解析;
為“神經(jīng)元興奮驅(qū)動(dòng)能量消耗”提供直接實(shí)驗(yàn)證據(jù);
推動(dòng)神經(jīng)代謝研究從宏觀向細(xì)胞尺度深化。
總結(jié)
本研究通過(guò)丹麥Unisense電極的高精度氧監(jiān)測(cè),結(jié)合光遺傳學(xué)與多模態(tài)技術(shù),首次闡明:
1.能量消耗的核心是神經(jīng)元興奮(電壓門(mén)控鈉通道活動(dòng)),而非網(wǎng)絡(luò)節(jié)律(如γ振蕩);
2.PV中間神經(jīng)元是代謝主動(dòng)單元,其抑制功能不額外增加氧耗;
3.Unisense技術(shù)的不可替代性:
揭示細(xì)胞類(lèi)型特異性氧代謝動(dòng)力學(xué);
為神經(jīng)能量分配理論提供定量依據(jù);
奠定精準(zhǔn)解析腦能量代謝的技術(shù)基石。
該研究重塑了對(duì)腦能量預(yù)算的基本認(rèn)知,Unisense電極在此過(guò)程中發(fā)揮了關(guān)鍵的機(jī)制解碼器作用。