熱線:021-66110810,66110819
手機:13564362870
熱線:021-66110810,66110819
手機:13564362870
Acquisition, Maintenance, and Ecological Roles of Kleptoplasts in Planoglabratella opercularis (Foraminifera, Rhizaria)
有孔蟲,Rhizaria 中盜賊質體的獲取、維護和生態作用
來源:Frontiers in Marine Science 1 July 2020 | Volume 7 | Article 585
摘要核心發現
本研究首次揭示潮間帶底棲有孔蟲Planoglabratella opercularis通過攝食硅藻獲取并維持其葉綠體(盜質體),形成獨特的混合營養策略。通過分子系統學、透射電鏡、氧氣微電極及細胞內pH成像等多技術整合,發現:
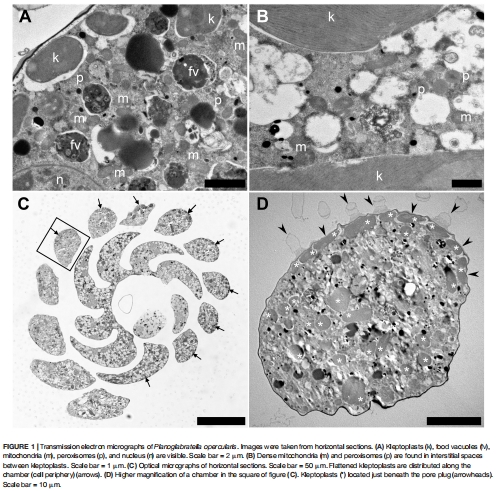
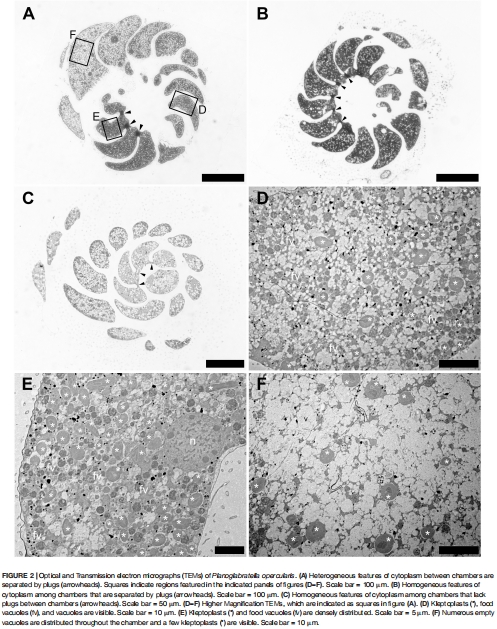
1. 盜質體源自附生硅藻(如Navicula和Nitzschia屬),定位在細胞外圍靠近孔塞處(圖1D),優化光捕獲;
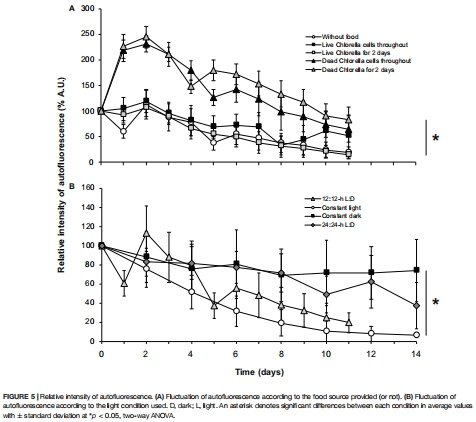
2. 盜質體壽命受食物供給與光照調控:無食物時11天內降解,黑暗或周期性光照可延長其活性(圖5);
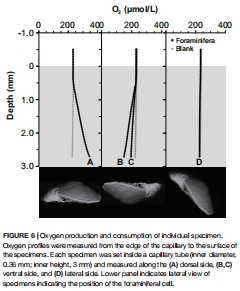
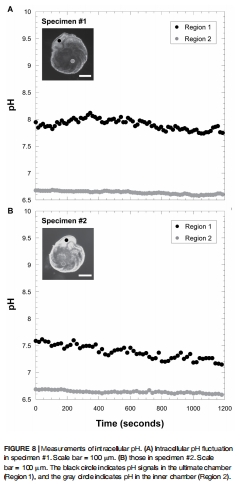
3. 盜質體光合作用產生氧氣(背側產氧率35.7 nmol O? hr?1 cm?2,圖6A),同時維持高細胞內pH(7.33-7.90,圖8),促進高鎂方解石殼體形成;
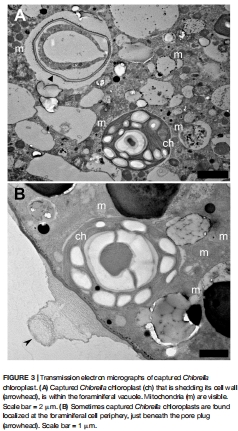
4.宿主可捕獲Chlorella葉綠體(圖3),拓展盜質體來源多樣性。
研究目的
1.盜質體生態機制解析:
闡明盜質體獲取途徑、維持條件及其在宿主能量獲取與鈣化中的作用。
2.代謝功能量化:
通過氧氣通量測量驗證盜質體光合活性,探索其對碳氮循環的貢獻。
3.環境適應性研究:
揭示光照、食物與鹽度梯度下盜質體功能的可塑性,解釋物種廣布性。
研究思路
1. 多尺度技術整合
形態定位:透射電鏡(TEM)觀察盜質體超微結構及分布(圖1-3);
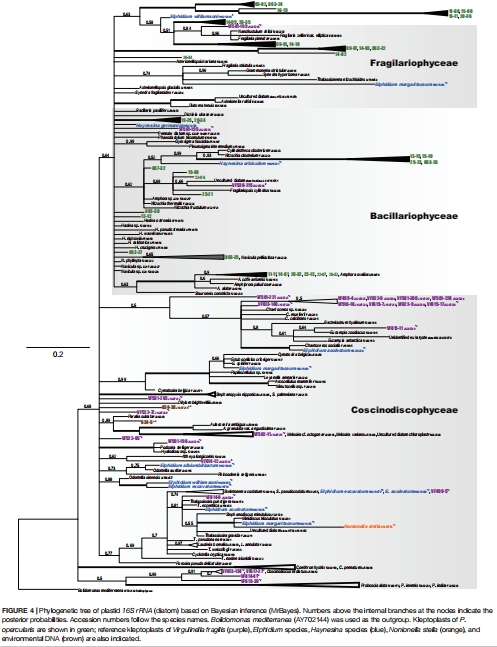
分子溯源:質體16S rRNA基因測序確定盜質體硅藻來源(圖4);
生理響應:
培養實驗結合自體熒光監測盜質體壽命(圖5);
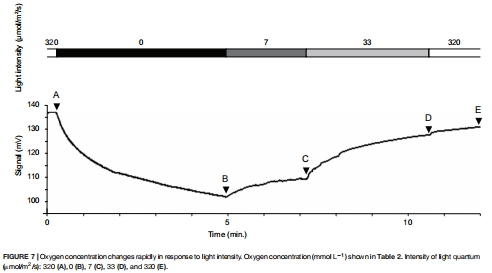
Unisense氧微電極測量背/腹側氧氣通量(圖6-7);
熒光探針(HPTS)成像細胞內pH梯度(圖8)。
2. 控制變量實驗設計
食物條件:活/死Chlorella投喂,對比盜質體維持效率;
光照梯度:全暗/全光/光暗周期(12:12-h、24:24-h),評估光合活性衰減;
跨種群采樣:日本四地潮間帶種群(Minami-Izu至Ofunato),驗證生態普適性。
關鍵數據及研究意義
1. 盜質體分布與形態(圖1-3)
數據:
盜質體密集分布于細胞背側近孔塞處(圖1C-D),呈扁平狀以最大化光捕獲;
Chlorella葉綠體可被捕獲并定位在相同區域(圖3B),拓展宿主營養策略。
意義:揭示空間優化策略——盜質體定位與氣體交換孔協同,提升光合效率。
2. 盜質體來源多樣性(圖4)
數據:
質體16S rRNA系統樹顯示盜質體源自多硅藻種(Navicula、Nitzschia等);
來源組成隨采樣點鹽度(25-34)與季節變化。
意義:證實宿主廣譜攝食適應性,支撐其在潮間帶多變環境中的生存。
3. 盜質體維持動態(圖5)
數據:
無食物時自體熒光11天內衰減81%(圖5A);
黑暗條件下熒光維持率74%(14天);
投喂熱滅活Chlorella可提升熒光強度245%(第2天)。
意義:光照與食物共同調控盜質體更新頻率,確立混合營養模式(自養+異養)。
4. 氧氣通量與pH調控(圖6-8)
數據:
背側光合產氧35.7 nmol O? hr?1 cm?2,腹側耗氧-22.9 nmol(圖6);
光照強度從320→0 μmol光子時,5分鐘內氧濃度從350→259 μmol/L(圖7);
終室pH 7.33-7.90 vs. 細胞中心pH 6.63-6.64(圖8)。
意義:盜質體驅動代謝極化(背側產氧/腹側耗氧),并維持堿性微環境促進鈣化。
丹麥Unisense電極數據的核心價值
技術突破性
單細胞分辨率:
20-30 μm微電極(OX-25)穿透毛細管(內徑0.36 mm),實現亞毫米尺度氧氣梯度解析(圖6-7);
動態過程捕捉:
50 μm步進式剖面+1秒/點采樣頻率,量化光照切換(320→0 μmol光子)下5分鐘級光合響應(圖7B)。
關鍵科學發現
1.盜質體光合活性量化:
背側產氧率35.7 nmol O? hr?1 cm?2(圖6A),證實盜質體貢獻≥50%宿主能量需求(對比全暗呼吸0.66 nmol O? h?1個體?1);
2.代謝空間分工:
腹側耗氧(-22.9 nmol)與偽足活動耦合(圖6B-C),揭示背光合成/腹側異養的功能分區;
3.光響應閾值:
氧產在33-320 μmol光子間無顯著差異(圖7D-E),暗示盜質體低光適應特性(與潮間帶弱光環境匹配)。
生態意義
碳泵新路徑:
盜質體光合固碳(79±33 mg C m?2 d?1)貢獻潮間帶碳循環,修正傳統異養模型;
酸化抵抗機制:
維持高pH(7.33-7.90)緩沖海水酸化對鈣化的抑制(圖8),為鈣質生物適應性提供新解;
混合營養模型:
氧氣通量數據支撐"光合+攝食"雙能源策略,解釋物種在寡營養潮間帶的競爭優勢。
結論
1.盜質體為多功能細胞器:
兼具能量生產(光合固碳)、鈣化調控(pH提升)、環境感應(光響應)三重角色;
2.生態可塑性基石:
盜質體來源多樣性(硅藻/Chlorella)與維持策略(光/食調控)支撐宿主廣鹽性分布;
3.方法學創新:
Unisense微電極首次實現有孔蟲單細胞代謝時空解析,為微型生物能量學研究樹立新標。
未來方向:量化海洋酸化下盜質體pH調控能力,整合組學解析宿主-盜質體基因轉移機制。
圖示關聯:
圖1:盜質體超微結構與定位
圖4:盜質體分子溯源
圖5:盜質體維持動態
圖6-7:Unisense氧氣通量測量
圖8:細胞內pH梯度