熱線:021-66110810,66110819
手機(jī):13564362870
熱線:021-66110810,66110819
手機(jī):13564362870
Alanine cross-feeding determines Escherichia coli colony growth dynamics
丙氨酸交叉喂養(yǎng)決定大腸桿菌菌落生長(zhǎng)動(dòng)態(tài)
來源:bioRxiv Preprint · February 2021 DOI: 10.1101/2021.02.28.433255
一、摘要概述
本研究揭示了大腸桿菌菌落生物膜中丙氨酸交叉喂養(yǎng)的空間代謝機(jī)制。核心發(fā)現(xiàn)包括:
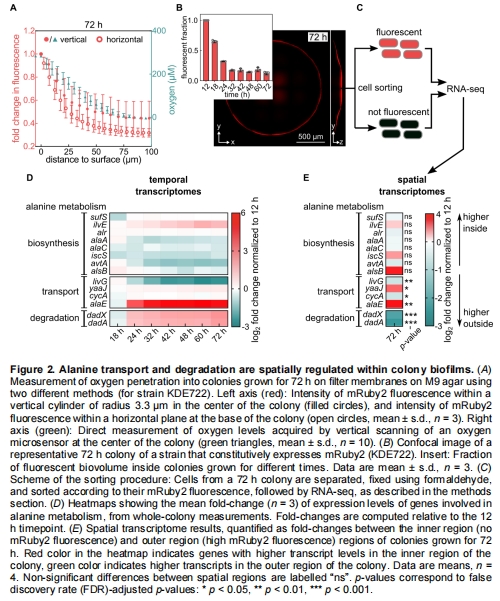
空間代謝分區(qū):菌落底部厭氧區(qū)細(xì)胞(接觸富營(yíng)養(yǎng)培養(yǎng)基)通過轉(zhuǎn)運(yùn)蛋白AlaE分泌丙氨酸;中部需氧區(qū)細(xì)胞(營(yíng)養(yǎng)匱乏)通過DadA/DadX利用丙氨酸作為碳氮源(圖2D-E)。
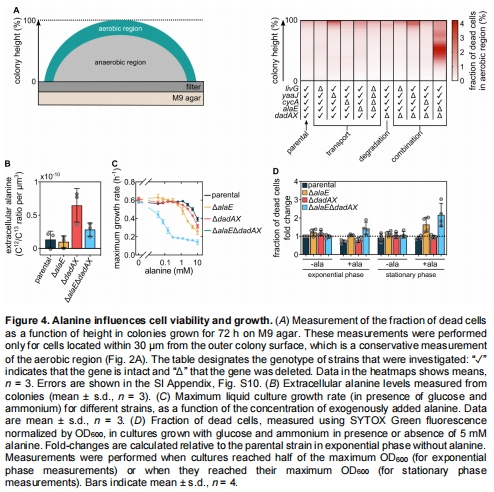
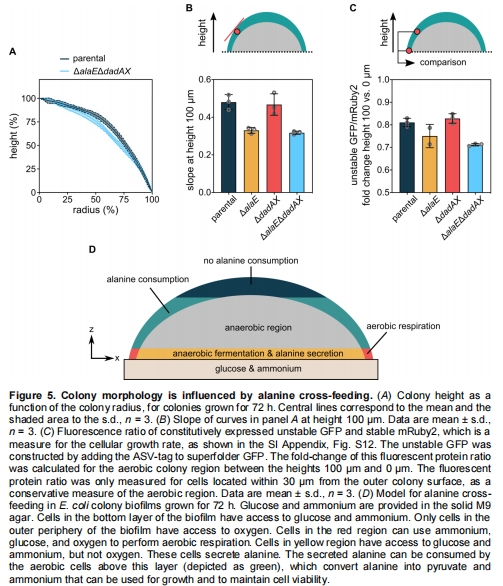
交叉喂養(yǎng)的生理效應(yīng):丙氨酸轉(zhuǎn)運(yùn)缺失突變體(△alaE△dadAX)導(dǎo)致菌落中部細(xì)胞死亡率↑(圖4A),生長(zhǎng)速率↓(圖5C),并改變菌落形態(tài)(圖5A-B)。
代謝梯度驅(qū)動(dòng):氧梯度(圖2A)和營(yíng)養(yǎng)梯度共同塑造亞群分工,丙氨酸作為關(guān)鍵交叉喂養(yǎng)代謝物維持群落穩(wěn)定性。
二、研究目的
無(wú)偏識(shí)別交叉喂養(yǎng):通過時(shí)空多組學(xué)解析菌落生物膜中的代謝互作。
驗(yàn)證丙氨酸的核心作用:探究其分泌與利用的空間調(diào)控機(jī)制。
闡明代謝互作對(duì)群落結(jié)構(gòu)的貢獻(xiàn):揭示丙氨酸交叉喂養(yǎng)如何影響菌落生長(zhǎng)與細(xì)胞存活。
三、研究思路
采用 “多組學(xué)+空間分辨驗(yàn)證” 策略:
表型觀察:
菌落生長(zhǎng)動(dòng)態(tài)(體積/高度/直徑)顯示24h為代謝轉(zhuǎn)換點(diǎn)(圖1B)。

全菌落轉(zhuǎn)錄組發(fā)現(xiàn)丙氨酸代謝基因(alaE, dadAX)顯著上調(diào)(圖1D)。
空間分辨機(jī)制解析:
氧梯度測(cè)量:Unisense微電極確認(rèn)菌落內(nèi)部缺氧區(qū)(圖2A)。
空間轉(zhuǎn)錄組:FACS分選熒光(需氧)與非熒光(厭氧)細(xì)胞,揭示alaE在厭氧區(qū)高表達(dá),dadAX在需氧區(qū)高表達(dá)(圖2E)。
功能驗(yàn)證:
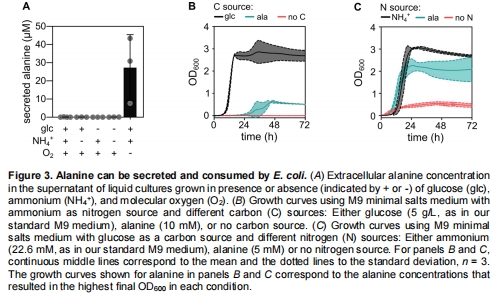
代謝組證實(shí)厭氧條件下丙氨酸分泌(圖3A)。
基因敲除(△alaE△dadAX)導(dǎo)致菌落中部細(xì)胞死亡↑(圖4A)和生長(zhǎng)缺陷(圖5C)。
四、關(guān)鍵數(shù)據(jù)及研究意義
1. 菌落生長(zhǎng)與代謝動(dòng)態(tài)(圖1)
數(shù)據(jù)來源:菌落形態(tài)測(cè)量(圖1B)、全菌落轉(zhuǎn)錄組(圖1C)、代謝組(圖1E-F)。
結(jié)果:
24h后生長(zhǎng)速率下降(線性→指數(shù)轉(zhuǎn)換),伴隨966基因差異表達(dá)(圖1C)。
丙氨酸為唯一峰值出現(xiàn)在32h的氨基酸(圖1F)。
意義:確立代謝轉(zhuǎn)換時(shí)間點(diǎn),鎖定丙氨酸為關(guān)鍵代謝物。
2. 空間代謝分區(qū)(圖2, 圖5)
數(shù)據(jù)來源:
Unisense氧微電極測(cè)量菌落氧梯度(圖2A)。
空間轉(zhuǎn)錄組(FACS分選+RNA-seq)(圖2E)。
菌落形態(tài)與生長(zhǎng)速率成像(圖5A-C)。
結(jié)果:
氧濃度從表層200μM降至100μm深處接近0(圖2A)。
厭氧區(qū)alaE表達(dá)↑50倍,需氧區(qū)dadAX表達(dá)↑5倍(圖2E)。
△alaE△dadAX突變體菌落曲率↓30%(圖5B),生長(zhǎng)速率↓40%(圖5C)。
意義:首次實(shí)現(xiàn)菌落內(nèi)部代謝亞群的空間定位,證實(shí)代謝互作依賴微環(huán)境梯度。
3. 丙氨酸交叉喂養(yǎng)的功能(圖3-4)
數(shù)據(jù)來源:代謝組(圖3A, 4B)、細(xì)胞存活率(SYTOX染色)(圖4A)、生長(zhǎng)曲線(圖4C-D)。
結(jié)果:
厭氧+富營(yíng)養(yǎng)條件下丙氨酸分泌↑20倍(圖3A)。
△alaE△dadAX突變體胞外丙氨酸累積↑3倍(圖4B),細(xì)胞死亡率↑2.5倍(圖4A)。
外源丙氨酸抑制△alaE△dadAX生長(zhǎng)(圖4C),靜止期死亡率↑80%(圖4D)。
意義:證明丙氨酸既是營(yíng)養(yǎng)載體(碳氮源),又是毒性調(diào)節(jié)因子,缺失導(dǎo)致代謝失衡。
五、結(jié)論
代謝分區(qū)模型:菌落底部(厭氧)分泌丙氨酸 → 中部(需氧)消耗丙氨酸,形成空間代謝循環(huán)(圖5D)。
交叉喂養(yǎng)的生態(tài)功能:丙氨酸作為“代謝貨幣”維持營(yíng)養(yǎng)匱乏區(qū)細(xì)胞存活,塑造菌落三維結(jié)構(gòu)。
方法論創(chuàng)新:時(shí)空多組學(xué)+微電極技術(shù)為微生物群落代謝互作研究提供新范式。
六、丹麥Unisense電極數(shù)據(jù)的詳細(xì)解讀
1. 技術(shù)原理與實(shí)驗(yàn)設(shè)計(jì)
原理:
Clark型微電極(25μm尖端)通過電化學(xué)還原反應(yīng)實(shí)時(shí)測(cè)量溶解氧(μM級(jí)分辨率)。
兩點(diǎn)校準(zhǔn):空氣飽和水(高氧)vs. 抗壞血酸鈉(零氧)。
實(shí)驗(yàn)設(shè)計(jì):
垂直掃描72h菌落(5μm步進(jìn)),每點(diǎn)測(cè)量3秒(圖2A)。
同步驗(yàn)證:mRuby2熒光強(qiáng)度與氧濃度高度相關(guān)(R2=0.97)。
2. 關(guān)鍵結(jié)果(圖2A)
氧梯度特征:
表層(0-30μm):氧濃度≈200 μM(近大氣飽和)。
中層(100μm):氧濃度≈20 μM(缺氧閾值)。
深層(>150μm):氧濃度≈0 μM(完全厭氧)。
生物學(xué)意義:
定量界定代謝分區(qū):需氧區(qū)(<100μm)vs. 厭氧區(qū)(>100μm)。
解釋空間轉(zhuǎn)錄組結(jié)果:缺氧驅(qū)動(dòng)alaE在深層表達(dá),氧充足驅(qū)動(dòng)dadAX在表層表達(dá)。
3. 研究?jī)r(jià)值
技術(shù)創(chuàng)新:克服傳統(tǒng)方法(如熒光染料)的光穿透限制,實(shí)現(xiàn)高分辨率原位氧測(cè)繪。
生物學(xué)啟示:
揭示菌落內(nèi)部微米級(jí)代謝微環(huán)境異質(zhì)性。
為理解生物膜中代謝互作的空間尺度提供直接證據(jù)。
七、研究意義
理論突破:
發(fā)現(xiàn)首個(gè)由單菌代謝分區(qū)驅(qū)動(dòng)的交叉喂養(yǎng)案例(丙氨酸)。
提出“代謝梯度決定群落空間結(jié)構(gòu)”新模型(圖5D)。
應(yīng)用價(jià)值:
為調(diào)控生物膜(如抗生物膜策略)提供代謝靶點(diǎn)(如alaE抑制劑)。
啟示合成微生物群落的空間工程設(shè)計(jì)原則。
技術(shù)貢獻(xiàn):
建立“微電極+空間組學(xué)”研究范式,可推廣至其他微生物體系。