熱線:021-66110810,66110819
手機:13564362870
熱線:021-66110810,66110819
手機:13564362870
Co-expression analysis reveals distinct alliances around two carbon fixation pathways in hydrothermal vent symbionts
共表達分析揭示了熱液噴口共生體中圍繞兩條碳固定途徑的不同聯盟
來源:Nature Microbiology | Volume 9 | June 2024 | 1526–1539
摘要核心內容
深海熱液管狀蠕蟲Riftia pachyptila的內共生細菌"Candidatus Endoriftia"同時具備兩種碳固定途徑——卡爾文-本森-巴斯漢(CBB)循環和還原性三羧酸(rTCA)循環。本研究通過高壓培養實驗結合轉錄組、代謝速率測量及共表達網絡分析,揭示兩種途徑在不同地球化學條件下的協調機制:rTCA與氫化酶和異化硝酸鹽還原協同,CBB與硫氧化和同化硝酸鹽還原協同。碳固定速率高達24μmol g?1h?1,表明雙途徑共存是維持高生產力的適應性策略。
研究目的
闡明兩種碳固定途徑(CBB和rTCA)在動態熱液環境中的表達調控關系。
探究兩種途徑與其他代謝過程(硫、氮、氫代謝)的功能協同性。
量化共生體的凈碳固定速率,評估雙途徑的生理意義。
研究思路
樣本采集與模擬培養:
從東太平洋海隆(EPR)采集Riftia pachyptila,在高壓呼吸儀(HPRS)中模擬原位溫壓和地球化學條件(圖1d-f)。
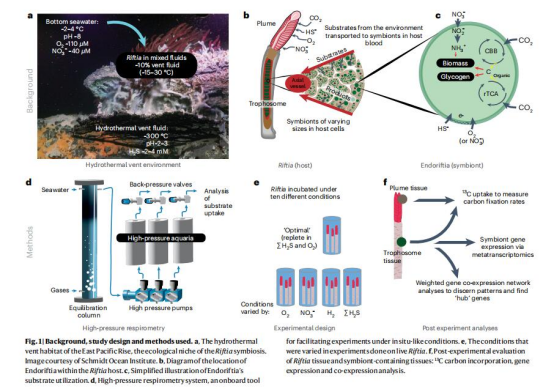
設計多組處理:富硫/氧(replete)、限硫/氧(limiting)、無底物對照(control)(圖1e)。
多維度數據測量:
底物攝取速率:ΣH?S、O?、H?(使用丹麥Unisense電極等)。
碳固定速率:13C標記DIC(溶解無機碳)摻入生物量速率(Cinc)。
轉錄組分析:共生體組織mRNA測序(RNA-seq),差異表達(DE)和共表達網絡(WGCNA)。
整合分析:
關聯環境參數-基因表達-代謝速率,識別關鍵響應模塊(如rTCA與氫化酶的協同)。
測量數據及其研究意義
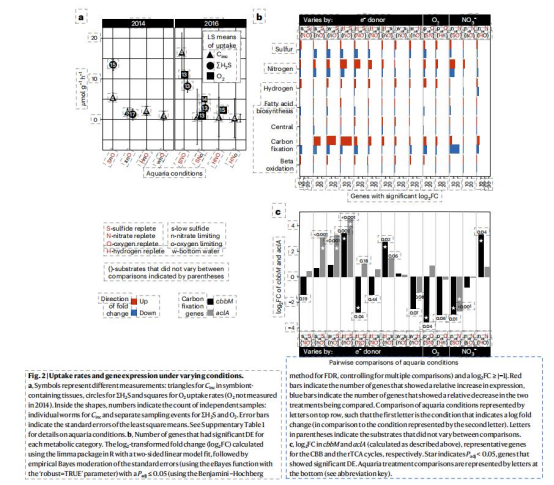
底物攝取速率(ΣH?S、O?、H?)
數據來源:Fig.2a
研究意義:
ΣH?S與O?攝取高度正相關,證實硫氧化需氧支持(經典能量代謝基礎)。
H?攝取率與O?無相關性,且與對照組無差異,排除氫氣作為主要電子供體的作用(修正傳統假設)。
碳固定速率(Cinc)
數據來源:Fig.2a
研究意義:
最高達24μmol g?1h?1,超過多數化能自養群落,與高光合生物相當,證明雙途徑驅動超高生產力。
限硫條件下Cinc顯著下降,凸顯硫對碳固定的核心限制作用。
基因表達數據
差異表達(DE):
數據來源:Fig.2b-c,Extended Data Fig.1
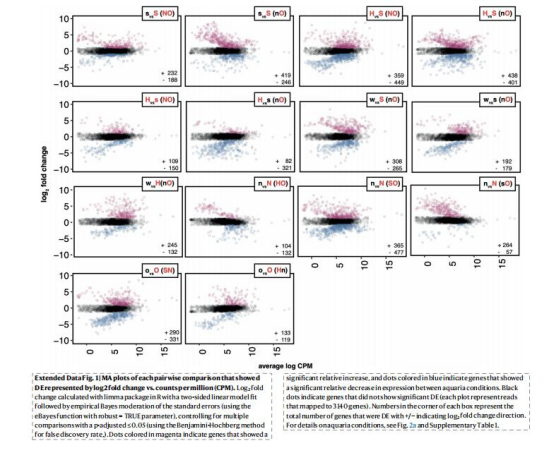
研究意義:
rTCA關鍵基因(aclA)在限硫時上調,CBB基因(cbbM)在缺氧時下調(Fig.2c),表明途徑對環境擾動的差異化響應。
硫代謝基因在限硫/氧時下調,氫化酶與氮代謝基因上調(Fig.2b),揭示能量短缺時的代謝重塑。
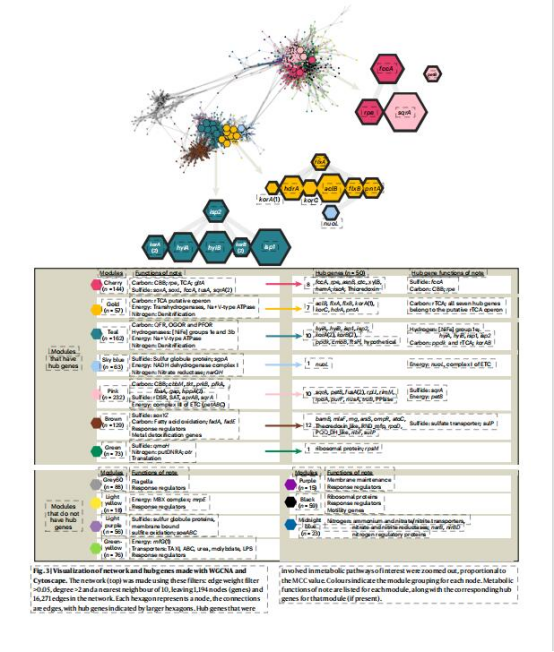
共表達網絡(WGCNA):
數據來源:Fig.3,5
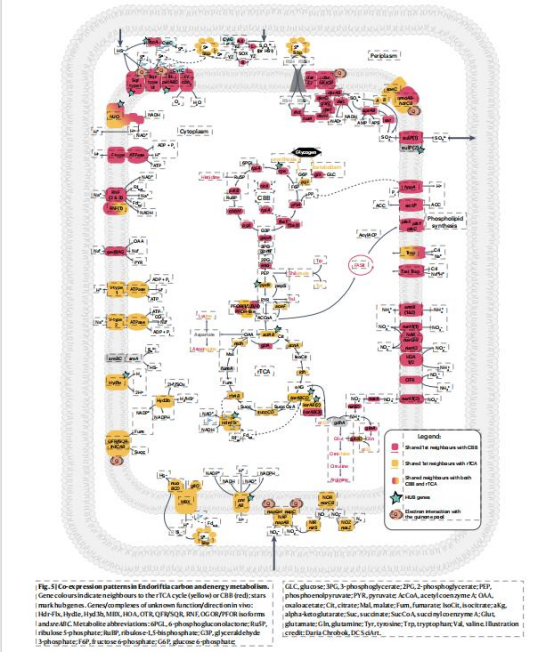
研究意義:
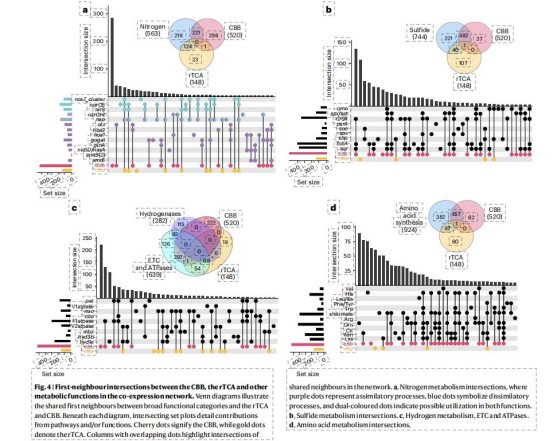
rTCA與氫化酶(Hyd1e)、異化硝酸鹽還原(norCB,nosZ)強關聯(金色/青綠色模塊);CBB與硫氧化(sox,fccA)、同化硝酸鹽還原(narGHI)協同(粉色/櫻桃色模塊)(Fig.5),揭示途徑特異性代謝聯盟。
僅1個基因與雙途徑共享鄰居,表明調控獨立性(Fig.4)。
丹麥Unisense電極數據的詳細解讀
測量目標:溶解H?濃度(使用Unisense H?minisensor 500,檢測限0.3μM,范圍0–800μM)。
關鍵結果:
H?攝取率在實驗組與對照組無顯著差異(Fig.2a),且與O?攝取無相關性(Supplementary Table 2)。
研究意義:
否定氫氣作為關鍵能源:推翻早期假設,明確ΣH?S(而非H?)是共生體主要電子供體。
解釋氫化酶功能矛盾:盡管氫化酶基因高表達且為網絡樞紐(Fig.3),但H?代謝不貢獻凈碳固定,暗示其可能參與:
胞內氫循環(如暗發酵中維持氧化還原平衡)。
硫還原耦合產ATP(通過psrABC/sreABC基因)。
深化代謝模型:為Fig.5中氫化酶(Hyd1e,Hyd3b)與rTCA的共表達提供生理背景,提示其在能量應激中的備用角色。
結論
雙途徑分工機制:
CBB主導富硫/氧區,驅動硫氧化耦合的高效碳固定;
rTCA在限硫/氧時激活,聯姻氫化酶與異化硝酸鹽還原維持代謝(Fig.5)。
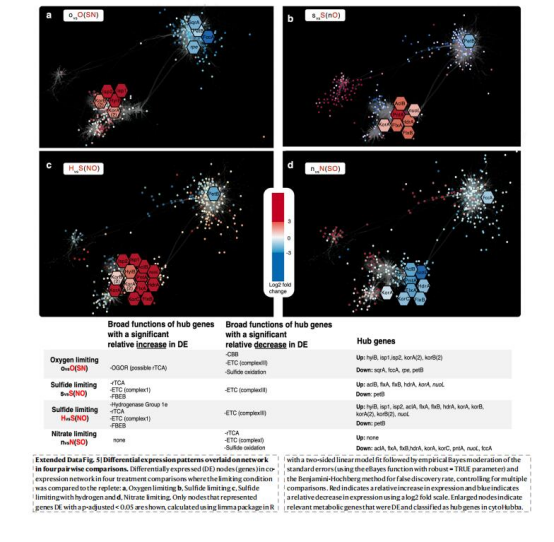
環境適應性:
雙途徑的模塊化調控(如限硫時rTCA-Hyd1e上調)增強對熱液動態環境的魯棒性(Extended Data Fig.5)。
生態意義:
超高碳固定速率(24μmol g?1h?1)是雙途徑共存的進化優勢,支撐管狀蠕蟲群落的高生物量。